Människan har sitt ursprung i Afrika. Storskalig bekräftelse av den "afrikanska" teorin om mänskligt ursprung har erhållits. kommentarer: Våra förfäder lämnade inte Afrika
Analys av kraniometriska (det vill säga relaterade till mätningar av skallen) för moderna människor indikerar att alla människor som lever på jorden idag härstammar från en relativt liten grupp individer som levde i Centralafrika för 60-80 tusen år sedan. När ättlingarna till dessa människor spreds runt om i världen förlorade de en del av sina gener och blev mindre och mindre mångfaldiga. I en artikel som nyligen publicerats i tidskriften Natur, hypotesen om ett enda ursprungscentrum för den moderna människan bekräftades av analysen av inte bara molekylärgenetiska data, utan också fenotypiska data (i detta fall storleken på skallen).
Mer och mer data som samlats in under de senaste åren tyder på att den "moderna" människan bildades i Ekvatorialafrika för 150-200 tusen år sedan. Dess spridning över planeten började för ungefär 60 tusen år sedan, när en relativt liten grupp människor flyttade till den arabiska halvön, och därifrån började deras ättlingar gradvis spridas över hela Eurasien (förflyttade sig främst österut längs Indiska oceanens kust), och sedan i hela Melanesien och Australien.
Processen för mänsklig bosättning på vår planet, enligt denna hypotes, borde ha åtföljts av en minskning av det ursprungliga lagret av genetisk variabilitet. När allt kommer omkring är det i varje skede inte hela ”föräldrarnas” population som ger sig av på sin resa, utan någon liten del av den, ett prov som omöjligt skulle kunna inkludera alla gener. Med andra ord borde det finnas en grundareffekt – en kraftig minskning av den totala genetiska mångfalden med bildandet av varje ny grupp av migranter. Följaktligen, när människor sprider sig, bör vi upptäcka det gradvisa försvinnandet av ett antal gener, utarmningen av den ursprungliga genpoolen. I verkligheten kan detta visa sig i en minskning av nivån av genetisk variabilitet, och ju längre från bosättningskällan, desto större grad. Om artens ursprungscentrum (i detta fall Homo sapiens) inte en utan flera, då blir bilden en helt annan.

Hypotesen om ett enda ursprungscentrum för moderna människor bekräftades nyligen av molekylärgenetiska data som samlats in som en del av det internationella Human Genome Diversity Project (HGDP). Den genetiska mångfalden i mänskliga populationer minskade med avståndet från Centralafrika, det förmodade centrum för mänskligt ursprung (se t.ex. Ramachandran et al. 2005). Det förblev dock oklart om denna effekt kunde upptäckas genom att hänvisa till fenotypiska egenskaper, till exempel de anatomiska egenskaperna hos moderna människor.
Andrea Manica från institutionen för zoologi vid University of Cambridge (UK) tillsammans med kollegor från institutionen för genetik vid samma universitet och institutionen för anatomi vid Saga Medical School (Japan) tog sig an lösningen på detta problem. Materialet baserades på skallmått (kraniometriska indikatorer) som samlats in över hela världen. Totalt 4 666 manliga skallar från 105 lokala populationer och ytterligare 1 579 honskallar från 39 populationer analyserades. Data om manliga skallar tas som grund eftersom de är mer representativa. Skallar äldre än 2 tusen år inkluderades inte i analysen för att undvika mätfel associerade med dåligt bevarande av gamla ben.
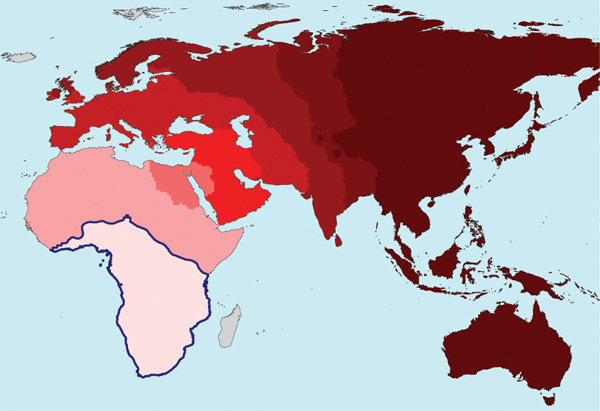
Resultaten av studien bekräftade hypotesen om ett enda centrum av mänskligt ursprung. Med avståndet från centrala Afrika minskade variabiliteten av skallens huvuddimensionella parametrar, vilket kan tolkas som en minskning av den initiala genetiska mångfalden. Ytterligare svårigheter med analysen var förknippade med det faktum att när människan bemästrade nya klimatzoner visade sig vissa egenskaper (eller visade sig inte vara) användbara och följaktligen stöddes eller inte stöddes av urval. Denna klimatanpassning påverkade också storleken på skallen, men användningen av speciella statistiska metoder gjorde det möjligt att isolera denna "klimatiska" komponent och inte ta hänsyn till den när man analyserade dynamiken i den initiala variabiliteten.

Parallellt, i samma arbete, bedömdes graden av genotyp heterozygositet för 54 lokala populationer av moderna människor. För detta ändamål använde vi data om mikrosatelliter (DNA-fragment som innehåller upprepningar), även insamlade som en del av HGDP-programmet. När de plottas på en karta visar dessa data en fördelning som är mycket lik den som avslöjas av fenotypiska egenskaper. När man rör sig bort från en persons ursprungscentrum minskar heterozygositeten (ett mått på genetisk mångfald), liksom den fenotypiska mångfalden.


Källa: Andrea Manica, William Amos, François Balloux, Tsunehiko Hanihara. Effekten av forntida befolkningsflaskhalsar på mänskliga fenotypiska variationer // Natur. 2007. V. 448. S. 346-348.
Se även:
1) Varför människan lämnade Afrika för 60 tusen år sedan, "Elements", 2006-06-30.
2) Mänsklighetens tidigaste historia reviderad, "Elements", 03/02/2006.
3) Mänsklighetens resa. Världens folk. Bradshaw Foundation (se fritt tillgänglig karta med animation som visar vägen för den tidiga människans spridning från Afrika).
4) Paul Mellars. Varför skingrade den moderna mänskliga befolkningen från Afrika ca. 60 000 år sedan. En ny modell (fulltext: Pdf, 1,66 Kb) // PNAS. 2006-06-20. V. 103. Nej. 25. s. 9381-9386.
5) Sohini Ramachandran, Omkar Deshpande, Charles C. Roseman, Noah A. Rosenberg, Marcus W. Feldman, L. Luca Cavalli-Sforza Stöd från förhållandet mellan genetiskt och geografiskt avstånd i mänskliga populationer för en seriegrundareffekt med ursprung i Afrika ( fulltext: Pdf, 539 Kb) // PNAS. 2005. V. 102. P. 15942-15947.
6) L. A. Zhivotovsky. Mikrosatellitvariabilitet i mänskliga populationer och metoder för att studera det // VOGiS Bulletin. 2006. T. 10. Nr 1. P. 74-96 (det finns en pdf av hela artikeln).
Alexey Gilyarov
Visa kommentarer (29)
Komprimera kommentarer (29)
Låt mig förklara populärt om genetisk drift. Anta att det finns någon stor population, till exempel 100 000 individer av en art (låt det vara en person, men med samma framgång kan det vara en vit hare, en luvtröja, en skogspelargon...). Om vi tar ett litet slumpmässigt urval av 10 individer från denna stora population, så kommer uppenbarligen inte alla gener som finns i föräldrapopulationen att hamna där, men de som gör det kommer, vid framgångsrik reproduktion och en ökning av storleken på dotterpopulationen, kommer att återges i många exemplar. Om man tar något annat litet prov från föräldrapopulationen parallellt, så kan andra gener råka komma dit, vilket också kommer att reproduceras i ett stort antal individer om någon ny population uppstår från detta prov. Följaktligen kan skillnader uppstå mellan sådana dotterpopulationer isolerade från varandra (vilket också kommer att visa sig i individers yttre utseende), som inte är resultatet av naturligt urval (d.v.s. inte adaptivt, inte adaptivt), utan erhålls helt enkelt p.g.a. någon slumpmässig kombination av omständigheter. Detta fenomen upptäcktes oberoende av Wright (som gav namnet "genetisk drift") och av våra landsmän, Dubinin och Romashov, som kallade det "genetisk-automatiska processer." Populationer av landlevande djur och växter från avlägsna oceaniska öar härstammar ofta från bokstavligen ett par individer. Naturligtvis är grundareffekten och genetisk drift särskilt uttalad i detta fall.
Mänsklig bosättning på den amerikanska kontinenten inträffade inte tidigare än för 25 tusen år sedan. Människor korsade där från den allra nordöstra delen av Asien längs "bron", ett stycke land (Beringia) som sedan förband Eurasien med Amerika. Då, för 18 tusen år sedan, fanns den sista starkaste glaciationen (is från norr nådde söderut till latitud 55) och den skar helt bort människor som flyttade till den amerikanska kontinenten (ättlingar till asiater) från kontakter med föräldrabefolkningen. Bildandet av indisk kultur började.
Alla främlingsfientliga och nationalister av alla slag (det spelar ingen roll om de föredrar den ariska rasen, negroider eller mongoloider) måste bli besvikna. Den moderna människan härstammar från en mycket liten grupp människor, där "Eva" är svart. Alla vi människor som lever på jorden är MYCKET NÄRA Släktingar. Till exempel är de genetiska skillnaderna mellan olika grupper av schimpanser som lever i olika områden i Centralafrika mycket mer betydande än skillnaderna mellan representanter för olika raser av Homo sapiens. Förlusten av genetisk (och, som visas i den diskuterade artikeln, fenotypisk) mångfald när vi flyttar bort från vårt gemensamma hemland - Afrika, är ytterligare ett kraftfullt bevis till förmån för hypotesen om ett enda ursprungscentrum för moderna människor. Liksom i fallet med människor finns utarmade genotyper till följd av befolkningens passage genom flaskhalsen (ett stadium med extremt låga antal) även hos andra djurgrupper. Till exempel, bland alla katter, upptar geparden en speciell plats. Alla geparder är också mycket nära släktingar, vilket inte kan sägas om lejon, tigrar, lodjur och huskatter. Jag ber om ursäkt för det omfattande, men jag hoppas att allt är klart nu.
Svar
Kära Alexey Gilyarov,
Det hände så att jag läste din anteckning och anteckningen "SENSATIONELLT FYND GÄLLER TEORIN OM "EXIDUS FRÅN AFRIKA"" (http://www.inauka.ru/evolution/article74070.html) i rad.
Där talar vi om upptäckten i Kina av ett skelett som är cirka 40 tusen år gammalt, som å ena sidan liknar en modern person, och å andra sidan är klart annorlunda än den afrikanska fenotypen.
Dessa data, enligt min åsikt, står i uppenbar motsägelse med materialet i din anteckning, och det skulle vara intressant att veta hur du kan lösa denna motsägelse.
Å andra sidan kan data om den afrikanska genotypens genetiska variation inte bara ha en "historisk" utan också en "biogeografisk" natur - till exempel kan man anta att afrikaner, I PRINCIPEN, på grund av vissa lokala geografiska eller klimatskäl, är mer aktiva finns det en process av genetiska mutationer som i synnerhet visar sig i fenotypisk mångfald. Om en sådan (ännu oupptäckt) process faktiskt äger rum, så bör i teorin tesen att den "mer mångfaldiga" afrikanska genotypen är en bekräftelse på afrikanernas "senioritet" korrigeras.
Personligen förefaller det mig som om tillståndet i teorin om mänskligt ursprung liknar situationen med taxonomin för kemiska grundämnen före tillkomsten av det periodiska systemet. Problemet var då att forskare försökte "naturligt" ordna alla KÄNDA data "i en rad", vilket inte lämnade något utrymme för de OKÄNDA, och DÄRFÖR fick de inget användbart. På samma sätt tyder förekomsten av motstridiga teorier om mänskligt ursprung, baserade på fast etablerade fakta, att VARJE av dessa teorier inte lämnar "luckor" för fakta som ÄNNU OKÄNDA - och därför är felaktiga.
Svar
Käre Mikhail, tyvärr, i anteckningen du hänvisar till anges varken källan (tidskriftens namn och artikelns koordinater) eller ens namnen på forskarna i engelsk transkription. Därför kan jag inte hitta originalpublikationen om det kinesiska fyndet som det hela började med, och det är helt enkelt omöjligt att bedöma utifrån en journalistisk text skriven utan någon förståelse för frågan. Så om du hittar koordinaterna för den ursprungliga (och inte sekundära) publikationen, rapportera det på webbplatsen! Det är troligt att detta inte alls är Homo sapiens, utan någon annan representant för hominiden. Om de tidigare i decennier talade om felande länkar i mänsklig paleontologi, nu finns det till och med ett överskott av dem. Alla stora antropologer är i alla fall överens om att det fanns en period på jorden då flera hominider SAMMANFATTAS på en gång, d.v.s. flera typer av forntida "människor" (citat - eftersom människor förstås i vid mening, inklusive till exempel neandertalare, som samexisterade med Homo sapiens i Europa under lång tid, men sedan dog ut). Så resterna av "förfäderna" är mestadels representanter för laterala linjer (som senare dog ut), och inte alls de verkliga förfäderna till Homo sapiens.
När det gäller antagandet om några särskilt höga mutationsfrekvenser hos afrikanska mänskliga förfäder, finns det ingen grund för det. Låt oss ändå följa Occams regel och inte skapa enheter utöver behovet.Svar
En tidig modern människa från Tianyuan Cave, Zhoukoudian, Kina
(Sen pleistocen | Neandertals | mandibel | postkranier | paleopatologi)Hong Shang*, Haowen Tong*, Shuangquan Zhang*, Fuyou Chen* och Erik Trinkaus
================Vad gäller Occams rakhyvel... Det här är en MYCKET bra teknik, men du måste använda den försiktigt, annars kan du skära av det som är helt klart nödvändigt :))
I exemplet med det periodiska systemet begick Mendeleev en mycket allvarlig "kränkning" av denna princip - och han visade sig ha rätt.
När jag jämför kartorna du tillhandahållit med kartorna över bosättningen Homo Sapiens (eller åtminstone med datumen för bosättningen i Asien och Europa), ser jag en uppenbar motsägelse. Om vi utgår från teorin om genetisk drift, så ju senare ett visst territorium befolkades, desto mindre genvariabilitet borde det finnas. Enligt tillgängliga data bosattes Europa senare än Asien, och borde därför vara "mörkare" än Asien. Eller mer generellt sett borde korten du tillhandahållit ha varit "fläckiga". Men på dem ser vi en "kontinuerlig gradient" - som om bosättningen från Afrika gick från söder till norr (Afrika-Europa) och sedan från väst till öst (Europa - Asien). Förvirrar inte sådana inkonsekvenser dig? Om dessa kartor visades för mig och ingen ytterligare förklaring gavs om vad som visades där, skulle jag där se en tydlig indikation på manifestationen av något planetariskt geofysiskt fenomen och skulle fråga hur situationen är i en annan del av världen (dvs. i Amerika).
Svar
Tack så mycket för länken. Tyvärr är det bara abstract som är öppet som man kan lära sig lite av. Jag ska försöka logga in från universitetsdatorn, kanske får jag hela texten. När det gäller dina kommentarer om bosättningen i Europa och Asien kan jag inte helt motivera författarens synpunkt. Du måste fråga dem detta. Titta på korten
som refereras på Elements (särskilt med animation!). Folk åkte till Europa ganska tidigt (men redan från Asien). Ja, och i PNAS finns helt öppna verk (om detta inte är det allra sista året). Det finns fortfarande inkonsekvenser, naturligtvis. Detta är inte förvånande, eftersom vi nyligen inte visste något alls. De kunskapsframsteg som bokstavligen har uppnåtts under de senaste 10-20 åren är överraskande.Svar
Jag hoppas få se en recension av den här artikeln i Elements.
Tack så mycket för den animerade kartan - det är precis vad jag letat efter länge.
Har du någonsin stött på kartor (statiska eller animerade) där arkeologiska bevis på människors tekniska framsteg (stenredskap, bostäder etc.) skulle ritas upp i kronologisk ordning? Eller kanske det finns resurser någonstans som skulle kunna användas för att bygga en sådan karta?
http://site/news/430144
Svar
Ja, jag läste den här artikeln på en gång. Tyvärr överensstämmer det inte riktigt med diskussionsämnet.
Den säger att teorin om förflyttning av de senaste mänskliga förfäderna (tredje expansionsvågen, för cirka 100 tusen år sedan) inte är sann, och genetiska data indikerar att biologiskt sett är vi människor ättlingar till alla invandrare från Afrika, med början för cirka 2 miljoner år sedan .
Om vi tar hänsyn till detta faktum (och jag ser ingen mening med att argumentera med det), så kan jag väl hålla med om påståendet att en grupp människor från Afrika bosatte sig i Kina för ett par miljoner år sedan, och vid tiden Homo Sapiens dök upp, de hade förändrats så mycket, vilket inte längre alls var som hennes afrikanska förfäder. Kanske var det denna grupp som gav upphov till synantroperna, och de i sin tur gav upphov till de moderna kineserna och asierna.
I själva verket, från min synvinkel, är frågan INTE om neandertalare kunde ha korsat sig med Cro-Magnons, eller om representanter för den 3:e vågen kunde ha korsat med företrädare för tidigare "expansionsvågor". Allt detta, ur min synvinkel, har INGEN betydelse i förhållande till problemet med sinnets utseende på jorden, eftersom det relaterar till kroppens utveckling, men inte medvetandet.
Men det som VERKLIGEN betyder något är att ta reda på orsakerna till KULTURBLASTEN.
Med "kulturell explosion" menar vi en SKÄRP tidsgräns (för ungefär 40-50 tusen år sedan), varefter människor började exponentiella framsteg inom teknik, kultur och miljöutveckling. Egentligen kan vi anta att Homo sapiens (d.v.s. den moderna bäraren av medvetande) dök upp exakt då - för ungefär 50 tusen år sedan, och inte för 150, och särskilt inte 800 tusen år sedan. Ur denna synvinkel har alla våra förfäder (inklusive representanterna för den tredje "expansionsvågen" som nämns överallt) som levde före denna "fatala punkt" ingenting gemensamt med oss när det gäller deras medvetandenivå, även om de är biologiskt "nästan identisk" med oss. Jag gav argument för detta antagande i en annan diskussion (se?discuss=430541). Och ingen analys av DNA från MODERNA människor kommer tyvärr att svara på orsakerna till denna "klyfta i medvetande".
Svar
: Med "kulturell explosion" menar vi en SKARP tidsgräns (för ungefär 40-50 tusen år sedan), varefter människor började exponentiella framsteg inom teknik, kultur och miljöutveckling.
Hur bedömdes det absoluta värdet av nivån på teknik, kultur och miljö? Finns det någonstans en illustration av en graf på vilken uppskattningar av denna nivå ritas baserat på kända fakta, och från vilken man skulle kunna dra en slutsats om den exponentiella tillväxten vid den tiden, och punkten för dess början, om det fanns någon? Finns det någonstans en analys av förändringar i miljöförhållanden eller andra faktorer som skulle kunna fungera som incitament för att höja denna nivå? Slutligen skulle det vara intressant att läsa vilka incitament som finns för att höja denna nivå nu. :-)
: Egentligen kan vi anta att Homo sapiens (d.v.s. den moderna bäraren av medvetande) dök upp exakt då - för ungefär 50 tusen år sedan, och inte för 150, och särskilt inte för 800 tusen år sedan. Ur denna synvinkel har alla våra förfäder (inklusive representanterna för den tredje "expansionsvågen" som nämns överallt) som levde före denna "fatala punkt" ingenting gemensamt med oss när det gäller deras medvetandenivå, även om de är biologiskt "nästan identisk" med oss. Jag gav argument för detta antagande i en annan diskussion (se?discuss=430541). Och ingen analys av DNA från MODERNA människor kommer tyvärr att svara på orsakerna till denna "klyfta i medvetande".
Svar
>Hur bedömdes det absoluta värdet av nivån på teknik, kultur och miljö?...
Läs diskussionen som jag länkade till. De frågor du tog upp diskuterades delvis där; jag presenterade i synnerhet en indirekt metod med vilken man kunde kvantifiera (d.v.s. få en visuell graf och inte generella resonemang). På det här diagrammet, om du ritar det, kommer "startpunkten" att vara ganska tydligt synlig.
När det gäller själva "kulturexplosionen" är detta ett ganska välkänt faktum. Det är bara det att efter denna tidsgräns blev verktygen mer eleganta och mer perfekta, teckningarna blev mer realistiska, vardagliga och kulturella föremål blev mer mångsidiga, och, viktigast av allt, under dessa 50 tusen år "fick" vi från en stenkniv till rymdskepp (detta gäller även frågan om utvecklingen av miljön). Och ALLA våra förfäder förbättrade under en liknande tidsperiod bara stenkniven något. Läs diskussionen – den svarar nog på de flesta frågor som först dyker upp.
> Finns det någonstans en analys av förändringar i miljöförhållanden eller andra faktorer som skulle kunna ge incitament att höja denna nivå?
I samma diskussion försökte jag visa att för det första måste dessa förhållanden vara MYCKET specifika (de måste nämligen innebära ett mycket strikt evolutionärt urval för graden av utveckling av medvetandet, som vi aldrig observerar i verklig levande natur), och, för det andra, under den aktuella tidsperioden (40-50 tusen år sedan) fanns det inga förhållanden på jorden alls som antydde en ökad artbildning. Det vill säga, baserat på logik och kända fakta BORDE det mänskliga sinnet helt enkelt INTE ha dykt upp på vår planet. Men det dök upp, och det får dig att undra över saknade fakta eller felaktiga antaganden som ligger till grund för den logiska analysen.
>> Och ingen analys av MODERNA människors DNA kommer tyvärr att svara på orsakerna till denna "klyfta i medvetande".
> För det första, försöker han verkligen svara på _den här_ frågan? Vad jag förstår så berör det honom inte alls.
Det är meningen, det "berör dig verkligen inte alls"! Men i litteraturen relaterad till problemet med människors uppkomst, finns det en ihållande ersättning av begrepp. Där sätts ett likhetstecken mellan biologisk evolution (d.v.s. OBSERVERADE förändringar i genotypen och fenotypen) och medvetandets evolution. Forskare vägrar helt enkelt att inse den grundläggande skillnaden mellan dessa fenomen.
> För det andra är det faktum att det inte visar något fundamentalt brott för exakt cirka 50 tusen år sedan redan en del av svaret på denna fråga. :-)
Detta är ett FÖR grovt verktyg för att användas för att hitta sådana skillnader. Det är som att mäta bakterier med en elevs linjal.
Och sedan, om uppkomsten av mänskligt medvetande var resultatet av någon liten modifiering av genomet, så kommer en analys av moderna människors DNA inte ALLS att visa när denna modifiering inträffade och om den inträffade i princip, eftersom det finns hos ALLA människor, och det är helt enkelt omöjligt att förstå att detta är just en modifiering av det "förmänskliga" genomet.
> Var inte övergången från bakteriekolonier till encelliga inte mindre av en bristning? Var inte övergången från encelliga till flercelliga organismer ett mindre avbrott? Och så vidare.
Dessa frågor är också mycket intressanta, men för det första relaterar de specifikt till BIOLOGISK evolution och för det andra har de en grundläggande skillnad från frågan om uppkomsten av medvetande, eftersom skedde mycket mer ”naturligt”, dvs. över ganska långa tidsperioder (miljoner år) och genom försök och misstag. Och dessutom var de inte förknippade med en sådan helt onödig sak för överlevnad som Reason.
Svar
Hur vågar folk arbeta med statistik... På Rysslands territorium (förutom kanten av Kamchatka, verkar det som) finns det inte ett enda staket av dödskallar, men sedan målar de djärvt över dess territorium till en mycket specifik tillfällig bosättningszon!
Svar
När man rör sig bort från en persons ursprungscentrum minskar heterozygositeten (ett mått på genetisk mångfald), liksom den fenotypiska mångfalden.
Med andra ord, ju längre från Afrika, desto stabilare är de heterozygota och fenotypiska egenskaperna, d.v.s. hela uppsättningen egenskaper genomgick ett längre och mer noggrant urval och urvalet blev stabilt, vilket betyder att i dessa regioner är människor äldre än i Afrika, där de fortfarande är väldigt, väldigt unga, och så förändras de varje år, som barn när de växer upp.
Och i Afrika levde människor, närmare bestämt, på en linje parallell med ekvatorn, ungefär på Nordafrikas latitud, där glaciärer periodvis drev dem. Därifrån återvände de sedan, inte alla, hem när vädret värmde upp. Det är därför fåglarna flyger för att häcka i norr, också hemma, precis som människor. I Kenya, där man har grävt så entusiastiskt sedan upptäckten av "Lucy", finns helt enkelt unika förhållanden i form av en förskjutning av kontinentalplattan. De gräver inte där de "tappade" den, utan under "lyktan". Alla dessa kvarlevor av "urgamla mänskliga förfäder" kan mycket väl inte ha något med oss att göra. Förresten, genetisk analys har redan slagit ut neandertalaren ur den darwinska flocken, men hur de nyligen tvingade honom på oss som halvbröder! Afrika, som mänsklighetens förfäders hem, valdes tydligen på grund av civilisationernas paritet och politisk korrekthet. Troligtvis fanns det flera Adams, "av samma typ." Sex grundläggande mutationer, av 200 kända idag, tros finnas hos alla människor på jorden. Indikerar detta bara en gemensam förfader eller indikerar det villkoren för deras ursprung som är gemensamma för alla? Och är dessa markörer för mutationer? Det är möjligt att detta verkligen är ett "registreringsblad", men vad och varför? Jag kan inte acceptera förklaringen att naturen skapat en värdelös zon, detta finns inte i dess traditioner. Kanske 6 matcher är registreringskoden för vårt "postkontor" - Earth? Ha ha!
Svar
Faktum är att om du tittar på kartorna som ingår i artikeln som diskuteras kan du tydligt se att "något händer" i den afrikanska regionen, och intensiteten av detta minskar när det rör sig bort från centrum (dvs. Afrika). Detta fenomen kan dock förklaras på flera sätt, och det enklaste av dem (i enlighet med Occams princip) är att det vid "epicentrum" finns något MODERNT geofysiskt fenomen som återspeglas i biologiska processer, i synnerhet i frekvensen av mutationer av mänskligt genom.
Denna hypotes kan lätt testas - det räcker att göra samma "tillfälliga skanning" av gener, inte bara hos människor, utan också i andra arter som levde i Afrika med honom och har ungefär samma distribution på planeten. Om en liknande bild observeras i dem betyder det att materian befinner sig i geofysiska processer, men om det bara är hos människor betyder det att antingen hypotesen är felaktig eller att ytterligare faktorer måste beaktas.
Å andra sidan, en molekylär klocka, även om den inte ger den exakta tidpunkten för uppkomsten av en mutation, oavsett om du gillar det eller inte, visar den SEKVENSEN av mutationer. De där. om i Afrika denna mutation ÄNDÅ inte existerar, men i Asien finns den REDAN, betyder det att mutationen uppträdde EFTER att denna art uppträdde i Asien, och det är svårt att argumentera här. Såvitt jag förstår var det att döma av SEKVENSEN av ett antal mutationer som vi kom fram till att vi härstammar från Afrika. Politisk korrekthet har ingenting med det att göra - grovt sett är det bara att räkna på fingrarna.
Personligen är det som retar mig i alla diskussioner om människans ursprung det faktum att samtalet förs uteslutande kring strukturen av skallen, skelettet eller kromosomerna, d.v.s. runt något som kan grävas upp, mätas, brytas ner och vägas. Det är som att bedöma en persons intelligens efter storleken och stilen på hans kläder. Mer än storlek 50 är rimligt, mindre är det inte. Det finns en bröstficka - en sapiens, nej - en apa.
Rimlighet är för det första ett INFORMATIONSFEL. Och förmågan att bearbeta information återspeglas INTE i skelettet, inte heller i skallstrukturen, inte heller i de _för närvarande kända_ egenskaperna hos genomstrukturen. Även om biologer redan har insett att den genetiska sekvensen i sig inte betyder någonting - det som är viktigt är HUR gener "samverkar" i en LEVANDE organisms drift, och man kan inte ens drömma om att bedöma detta utifrån fossilt DNA. Så för tillfället är inte hela intelligensens "genetiska historia" värd ett öre. Det ger bara en ganska grov bild av vem som kom till denna värld efter vem.
Om vi bedömer uppkomsten av denna INFORMATIONSFÖRMÅGA (intelligens) hos människor efter det ENDA pålitliga (men tyvärr indirekta) materiella tecknet - föremål för materiell kultur, verktyg och hällmålningar, så visar det sig att intelligens uppstod SAMTIDIGT över hela planeten för ungefär 40 år sedan. För 50 tusen år sedan, dvs. bland ALLA människor som vid den tiden var bosatta över ett område på tusentals kilometer från Afrika till Australien. Om vi erkänner detta faktum, går alla "vetenskapliga" teorier om människors utseende omedelbart i avloppet, och vi står inför ett mycket obehagligt val - ingripandet av "högre makter" eller främmande intelligens.?diskutera= 430541), föreslog jag en "rimlig kompromiss" - "slumpmässig "viral introduktion av "sinnegener", men det ser inte heller särskilt övertygande ut. Även om detta ur min synvinkel är det bästa som kan erbjudas för tillfället, om man bestämt håller sig till den materialistiska synvinkeln.
Svar
Det stämmer, räkningen är bara på fingrarna, mer exakt på punktmutationer i Y-kromosomens icke-gena zon. Men det finns en poäng! Om vi tar, säg, Egypten, Mellanöstern eller Sydeuropa som den villkorliga ursprungspunkten för den "äldsta mutationen" - M168, så är den strategiska planen för beslagtagandet av planeten Jorden av progressiv mänsklighet i form av pilar på kartan är ritad lika korrekt. Faktum är till exempel att 10-15 % av icke-afrikaner inte har M89 (arabisk) mutator. Och om vi tar "utvandringen" genom Röda havet till den arabiska halvön som grund, då borde alla ha denna "snipp". Den genetiska databasen vid tidpunkten för studien innehöll endast cirka 50 tusen data, från, som du förstår, 3 miljarder män på jorden. Är detta ett tillräckligt prov? Vet inte. Jag tror nej. Men det visar redan att versionen av den tusenåriga simningen över Röda havet inte är korrekt. De australiska aboriginerna har den sista mutationen M9, dvs. i nästan 40 tusen år fanns det helt enkelt inga andra. Indianerna har också M3 och det är också tyst. Hur kan rörelsevägen i tid dras från antagandet - ett klipp per 5 tusen år. Alla dessa studier utförs endast i USA. USA är en ideolog inom globalismen. Globalismens viktigaste princip är "alla människor är bröder." Det är också viktigt att det inte finns någon äldre bland dem. De enda platserna som är mer idealiska än Afrika skulle vara Australien, Antarktis och Atlantis. Men det kommer inte att passa. Vem föreslog idén om att placera människans förfäders hemland i Afrika? Ja, fortfarande samma Mr. Darwin. "Monofilist", för helvete. Neandertalmänniskan (Nomo sapiens) ingick i den moderna människans linjära utvecklingskedja (Nomo sapiens sapiens) med rättigheterna, generellt sett, av en stamfader. Detta spelades in i Bol.Sov.Enz. svart, för helvete, "på ryska".
Svar
Kära Mikhail! Det finns ingen entydig bild av bosättning baserad på studier av mutationer. Med samma framgång kan startkontrollpunkten placeras till exempel i Spanien eller Egypten, eller till och med Mellanöstern. Bilden blir densamma. En "relativt liten grupp individer" korsar Gibraltar in i Afrika och drar sig tillbaka före glaciären. Den får en grundläggande mutation och delar sig sedan i en sydlig migration, längs Afrikas västkust, och periodvis "delas av", säg, längs floder, djupt in på kontinenten. Och österut - längs Medelhavskusten till Egypten, där den återigen delar sig i den sydafrikanska, migrerar uppför Nilen och Mellanöstern. Fram till denna punkt har alla samma mutationer. Sedan går en del till Mellanöstern (M89-mutationen saknas), och den andra delen, som snurrar runt den arabiska halvön, tar emot den. Du kan fortsätta som planerat idag. Bilden av mutationer är densamma. Vi måste också ta hänsyn till globala historiska processer. Erövringar av Makedonien, Rom, Arab och korståg, Mongol och andra. De kunde mycket allvarligt korrigera mönstret för nedärvning av mutationer i den manliga linjen. Det finns många andra punkter och oklarheter. Punktmutationer (snips) registreras strikt sekventiellt eller kan inträffa inom ett intervall (retrospektivt). Till exempel upprepningar av markörer i den sk. haplotyper kan förändras i vilken riktning som helst. Vad är karaktären på "klipp"? Varför uppstår de? Vad, slutligen, registreras i den icke-gena zonen av Y-kromosomen, vilken information? Det är trots allt inspelat och presenterat ganska strikt med mindre men stabila korrigeringar. Generellt sett är det för tidigt att göra globala generaliseringar.
Jag skulle vilja notera ytterligare en intressant punkt i förbigående. Det visar sig att slaviska haplotyper inte har mongoliska källor. Med tanke på att Y-kromosomen tydligt överförs genom den manliga linjen på ett ände-till-ände sätt, betyder detta att det inte finns några mongoler bland de slaviska förfäderna (inom ett rimligt tidsintervall). Så, "oavsett hur mycket ryska du skrapar, kommer du inte att hitta en mongol." Vilken gåva till Fomenko, som bevisar, om jag förstår honom rätt, att det mongoliska oket är en fiktion! Roligt, inte sant?Svar
Kära Vagant,
Jag förstår inte riktigt den ökade uppmärksamheten på genetik i historisk forskning. Tja, vi fick reda på att Djingis Khan försökte sitt bästa och i dag finns det 2 miljoner av hans ättlingar som springer runt i världen, så vad sägs om detta? Kanske en rad i Guinness rekordbok, ett märkligt faktum, men inget mer. Och vad gäller slaverna och mongolerna - kanske de faktiskt lyckades ta prover från dem vars förfäder inte korsade sig med mongol-tatarerna. Återigen, så vad? Upphäver detta historiska krönikor och utgrävningsresultat? Ett intressant tillägg till befintlig data, och inget mer. Det är fullt möjligt att tatarerna helt enkelt tog "sina" barn till horden, och därför borde vi inte leta efter mongoliska gener bland slaverna, utan slaviska gener bland ättlingarna till horden. Det visar sig vara en rolig slogan - "Ryssland är tatarernas hemland!" :) Men personligen är dessa "genetiska utgrävningar" helt ointressanta för mig.
Men det som verkligen är intressant är mysteriet med Reasons utseende på vår planet. Och här är frågan om huruvida intelligens först dök upp på ett ställe och därifrån spreds över planeten, eller oberoende - på flera ställen, fundamentalt viktig, även ur genetisk synvinkel.
Om intelligensens bärare endast dök upp på ett ställe (teorin om monocentrism), så tillåter detta oss att förklara varför alla människor representerar en biologisk art och har ungefär samma medvetandenivå. Samtidigt spelar det ingen roll var exakt det dök upp för första gången och vilka vägar det expanderade. Men denna teori förklarar inte hur mongoloiderna och kaukasierna uppträdde, eftersom det inte finns några bevis för omvandlingen av afrikaner till dessa raser (det finns inga övergångsformer). Dessutom stödjer inte arkeologiska bevis "erövringen" av Asien och Europa av afrikaner. Men samma problem uppstår om vi accepterar att sinnet uppstod i något annat, men bara centrum.
Om polycentristerna har rätt, och intelligens dök upp på flera ställen på basis av "lokalbefolkningen" (och det är precis vad som bekräftas av arkeologiska data!), så är det helt obegripligt hur varelserna, klart olika i genotyp, som gav upphov till folken i Afrika, Asien och Europa, lyckades förvandlas till samma art. Och det är ännu mer oklart vad som kunde ha orsakat en sådan omvandling. Detta motsäger i grunden allt som är känt inom genetiken idag. Men det vi vet kanske inte är allt som verkligen finns?
Därtill kommer problemet med rum-tid. Att döma av arkeologiska data inträffade omvandlingen av Homo Sapiens till Homo Sapiens Sapiens för cirka 50 tusen år sedan. En pålitlig indikator på denna omvandling är den "kulturella explosionen" - en förändring av hushållsartiklar, verktyg och uppkomsten av målning och konst. Människor vid den tiden ockuperade ett stort territorium - från Afrika till Australien. Och uppenbarligen skedde denna omvandling nästan omedelbart - under flera tusen år. Vilken typ av Djingis Khan var tvungen att gå längs kusten för att alla samtidigt skulle ha "medvetandegener"?
Således har vi idag situationen "Var du än kastar den, det finns en kil överallt." Och det genetiska sökandet efter det "historiska hemlandet" strävar bara efter ett mål - i inget fall att låta allmänheten tänka på problemen som nämns ovan. När allt kommer omkring, om en lösning "hittas", kan du förklara att alla problem har försvunnit och helt enkelt ignorera deras existens. Istället för ett smärtsamt sökande efter svar på svåra frågor finns det en länk till "de senaste vetenskapliga uppgifterna", som, trots deras noggrannhet, faktiskt inte bevisar eller förklarar någonting.
Svar
Kära Mikahail! Du ökade till och med ribban till 50 tusen år. Jag minns att jag fick lära mig att detta hände för 35-40 tusen år sedan. Men det är inte meningen. Det är viktigt att någon form av abrupt "reinkarnation" verkligen hände eller något. Vem (eller vad?) kom då ut ur Afrika för 80 tusen år sedan? Vad ska jag kalla honom? Det är klart att detta inte är Homo sapiens sapiens än, men det måste finnas någon sorts neoantrop. Om det här inte är en neandertalare, vem då? Inget svar! Genetiker säger att det inte är vår sak. Men det finns helt enkelt inga platser för andra neoantroper i åldern 80-100 tusen år. Den allmänna "Eva" tillskrivs i allmänhet 140-160 tusen år. Vem är hon då? Hon och "Adam" kan para sig, eftersom det finns en "vanlig" avkomma, vilket betyder att de är en art. Men detta är redan närmare skärningspunkten med de sista arkantroperna. Är det möjligt att mutationerna som studeras, gemensamma för alla, är de "vippbrytare" som satte igång sinnet och uppstod som ett resultat av en planetomfattande katastrof, oavsett bostadsort och ursprung? Det finns fortfarande fler frågor för genetiker än svar. En hypotes är bara en hypotes. Det är bara det att de "promoverar" det för mycket.
Svar
För mig personligen råder det inget tvivel om att varje levande organism (i grova drag, kapabla att reproducera sig självständigt) är en "mottagare" av ett eller annat "subtila fält", som västerländsk vetenskap inte vet något om än så länge. Enligt min åsikt står vi precis på tröskeln att öppna dessa fält. Kanske kommer de att kunna upptäckas och beskrivas med instrument om ytterligare 100-200 år. Men för nu, för "ortodoxa vetenskapsmän" är de ett strikt tabu - som allt som inte kan inkluderas i det befintliga vetenskapliga paradigmet.
Faktum är att det finns mer än tillräckligt med bevis för att biologiska organismer - från encelliga organismer till människor - ständigt "lyssnar" på sin yttre miljö. Det mest intressanta och övertygande argumentet för detta är behandling av sjukdomar med mycket svag millimeterstrålning (några till tiotals mikrowatt per kvadratcentimeter), som inte har NÅGON termisk effekt på vävnader och dessutom har en tydlig resonans. karaktär. Teorin om denna effekt har ännu inte konstruerats, även om själva effekten har varit känd i nästan 30 år och tusentals människor har botats med denna metod. Jag pratade om detta för att visa att levande varelser har mycket komplexa mekanismer som fungerar på molekylärgenetisk nivå, som är ansvariga för "uppfattningen" av strålning som kommer från det omgivande rummet. Dessutom är dessa mekanismer så känsliga och selektiva att de kan ta emot signaler som är mycket lägre än nivån av termiskt brus (vilket också är nonsens för ortodoxa fysiker som inte är bekanta med de levande systemens krångligheter). Och härifrån är det redan ett stenkast till att "ta emot" signaler som fortfarande är okända ultrasvaga, och därför inte mätta av hårdvara, fält.
Svar
Skriv en kommentar
Mänskligt ursprung- ett av de mest spännande och spännande ämnena inom vetenskap, filosofi och världsbild. Och en av de mest förvirrande. Faktum är att det inte finns ett enda direkt experiment som bestämt och entydigt skulle svara på frågan om var på planeten och när vår direkta förfader först dök upp, vilket skulle falla under den antropologiska beskrivningen av arten Homo sapiens och/eller "anatomiskt modern människa" (AMH). Här är varje koncept inte fixerat och är i huvudsak "flytande". Forntida skelettlämningar har hittats, men hur vet man om det är "första gången" eller om något ännu äldre kommer att hittas i morgon? Hur tillförlitliga är dateringar som faktiskt inte alls är tillförlitliga och som nästan alltid är omtvistade? Det finns dussintals antropologiska egenskaper som på något sätt prövas på konceptet Homo sapiens och på begreppet "anatomiskt modern människa", men teori är en sak (även om det fortfarande inte finns någon allmänt accepterad fullständig klassificering), och i praktiken är dessa egenskaper nästan omöjliga att tillämpa fullt ut - vanligtvis hittas bara fragment av skelettet, ofta utan ansiktsben, och för de äldsta benrester visar nästan alltid några "åldriga" drag.
Och då kommer det som kallas en vetenskapsmans samvetsgrannhet in. Insatserna är höga - varje nytt skelett eller dess fragment, vilket gör det möjligt att utropa det till "det äldsta kända" Homo sapiens eller ASP blir en världsomspännande sensation, med alla efterföljande konsekvenser i form av vetenskapliga utmärkelser, stora ekonomiska anslag, val till prestigefyllda vetenskapsakademier. Därför är tyvärr förvrängningar av data som beskrivs i akademisk och annan press, för att inte tala om populära publikationer, så angelägna om sensationer, så vanliga. Dejting i vetenskapliga publikationer är ibland uppblåst, ålderdomliga egenskaper "smetas över" och det blir mycket svårt att ta reda på var den verkliga informationen finns och var författarnas fantasier finns. Tvärsnittsstudier behövs, vilket är sällsynt. Slutligen finns det en hel del helt enkelt okvalificerat arbete, särskilt inom området populationsgenetik, eller arbete fokuserat på ett förutbestämt resultat.
Det är vad vår berättelse kommer att handla om. Nämligen om hur svårt det är att bryta igenom muren av partisk forskning, som är "fokuserad" på den förmodade uppkomsten av "anatomiskt moderna människan" förment från Afrika, och vad forskning faktiskt visar, ofta av samma författare, men tolkad i ett unikt sätt. Muren är också cementerad av ideologiska överväganden, för vilka det är nödvändigt att visa "Människans afrikanska ursprung", och den som hittar olika data och gör olika tolkningar är "rasist". Muren stärks av det faktum att nästan alla artiklar av engagerade forskare, och detta är majoriteten av populationsgenetiker, börjar med frasen " Som bekant kom den anatomiskt moderna människan ut från Afrika" Det vill säga att installationen pågår från allra första början. Detta ökar dramatiskt sannolikheten för att artikeln publiceras i en akademisk tidskrift.
Här är några exempel hämtade från titlarna på akademiska artiklar, eller från de första meningarna i artikelintroduktioner:
Human Origins: Out of Africa (artikeltitel; Tattersal, 2009);
Human Evolution and Out of Africa (från artikelns titel; Stewart och Stringer, 2012);
Afrikanskt ursprung för manlig (genetisk) mångfald (från artikelns titel; Cruciani et al, 2011);
Afrikanska ursprung för moderna östasiatiska människor (från artikelns titel; Ke et al, 2001);
...anatomiskt moderna människor anlände till Europa från Afrika för minst 45 tusen år sedan, efter spridningen av människor ut ur Afrika (Moorjani et al, 2011);
Moderna människor tros ha sitt ursprung i östra Afrika (Henn et al, 2011);
Det är allmänt accepterat att anatomiskt moderna människor har sitt ursprung i Afrika (Hammer et al, 2011);
Afrika, alla moderna människors förfäders hem (Lachance et al, 2012);
...avvikelsen mellan anatomiskt moderna människor från Afrika var ungefär 44 tusen år sedan (Underhill et al, 2000);
Moderna människor har sitt ursprung i Afrika för cirka 200 tusen år sedan (Campbell och Tishkoff, 2010);
... Anatomiskt moderna människor härstammar från en liten isolerad befolkning i Afrika för 150-200 tusen år sedan (Patin et al, 2009);
Sub-Sahara och nordöstra Afrika är de mest sannolika regionerna av mänskligt ursprung och en korridor till resten av världen (Arredi et al, 2004);
… mänsklig divergens började i Afrika (Ramachandran et al., 2005).
Nedan i detta arbete kommer det att visas att alla dessa bestämmelser, och liknande sådana, som återges i tiotals och hundratals akademiska och andra artiklar, är felaktiga.
Frågan är: hur nådde vetenskapen om mänskligt ursprung ett sådant liv? Hur kunde en ”expertkonsensus” ha utvecklats, baserad på en ensidig och sliten tolkning för ett förutbestämt svar? Hur kan en situation uppstå inom vetenskapen när andra rimliga tolkningar av samma eller andra data möts av uttryckt aggression, politiska anklagelser och oförställda negativa känslor? Varför blev "ut ur Afrika" en religion baserad på tro som inte behöver bevis?
Fram till 1980-talet var diskussionerna om mänskligt afrikanskt ursprung långsamma och i stort sett marginella. Två omständigheter hindrade oss allvarligt från att överväga detta. Först insåg man att den moderna människans avlägsna förfader var Homo erectus, Homo erectus, som uppstod för flera miljoner år sedan, möjligen i Afrika, men som var känt för att ha spridit sig över hela Eurasien för nästan två miljoner år sedan. Det är därför Homo sapiens, Homo sapiens, kunde bli hans ättling var som helst. För det andra visades det att ASP:s närmaste släkting, neandertalaren, inte bodde i Afrika. Därför, den gemensamma förfadern till den moderna människan och neandertalmannen, som levde, enligt olika källor, för mellan 600 och 300 tusen år sedan, visar det sig att han inte heller bodde i Afrika. Dessutom hade neandertalaren ljus hud, och vi kommer att uppehålla oss vid detta nedan. Därför kräver den moderna människans afrikanska ursprung ankomsten av en ljushyad direkt förfader till människan i Afrika, säg för 500-300 tusen år sedan, då var det hans oberoende, evolutionära förvärv av svart hud, annars skulle han inte överleva i Afrika, och sedan hans utträde från Afrika och hans oberoende förvandling tillbaka till svart ljus hud. På denna punkt uppfanns till och med en genialisk hypotes om D-vitaminets roll i den oberoende (utan korsning med ljushyade människor, som inte kunde existera utanför Afrika, annars skulle konceptet kollapsa) omvandlingen av svarta till ljushyade. , men denna hypotes bekräftades aldrig experimentellt. Det förblev spekulativt.
I allmänhet, fram till mitten av 1980-talet, var det inte särskilt allvarligt att prata om den moderna människans afrikanska ursprung. Men behovet av detta "i vissa vetenskapliga kretsar" eller, närmare bestämt, bland forskare med en viss liberal världsbild växte uppenbarligen, annars kunde den efterföljande utvecklingen av händelser inte förklaras. Det som hände var att 1987 publicerade tidskriften Nature en artikel av Rebecca Kann och medförfattare från University of California, Berkeley, med titeln "Mitochondrial DNA and Human Evolution." Artikeln är mycket svag, inte bara enligt moderna kriterier, utan också av den tidens, och man kan bara undra hur artikeln klarade recensionerna. Det räcker med att nämna att i sammanfattningen som föregick artikeln, rapporterades det att mitokondriella DNA som studerats av författarna härstammar från en enda kvinna som "postulerades" (!) för att ha levt för ungefär 200 tusen år sedan, "förmodligen" (! ) i Afrika.
Efter publiceringen av artikeln öppnade sig himlens avgrund, slussarna och portarna slogs upp. Den västerländska pressens entusiasm över att afrikaner är våra förfäder var fantastisk. Denna forntida afrikanska kvinna döptes omedelbart till Eva, och världens ledande tidningar publicerade information om detta på glansiga omslag. Sedan dess har den ihärdiga manipulationen av den allmänna opinionen fortsatt utan uppehåll, om inte ökat. Detta har blivit en allmänt accepterad åsikt, utmanande som är besläktad med att hävda möjligheten av en evighetsmaskin. Utmanaren går med andra ord emot den vetenskapliga ”konsensus”, som naturligtvis inte finns, men som ständigt förkunnas. Jag diskuterade detta problem med antropologer (som chefredaktör för den internationella tidskriften "Framsteg inom antropologi") har fått och fortsätter att få en hel del brev där professionella vetenskapsmän delar att de naturligtvis tvivlar eller kategoriskt inte håller med om att "människans afrikanska ursprung" åtminstone på något sätt är berättigat, men de vill inte tala om det i tryck, eftersom "dyrare för dig själv." Och för att en artikel i en vetenskaplig tidskrift fortfarande kommer att avvisas, oavsett vilken data som finns och hur den underbyggs.
Så vad står det i Rebecca Canns artikel från 1987? Vad låg till grund för den nya religionen? Hur började det hela? Låt oss ta en titt.
Kann et al.s (1987) framstående artikel om "ut ur Afrika"
I inledningen till artikeln finns inte ett ord om Afrika och mänsklighetens förmodade ursprung där. Det vill säga, artikeln positionerar sig som den första i detta avseende. Den experimentella delen av artikeln är bestämning av nukleotidsekvensen för mtDNA från 147 kvinnor från fem huvudregioner:
Afrika– 20 personer (två är födda söder om Sahara, resten är svarta invånare i USA, vanligtvis mestiser med en inblandning av Y-DNA från kaukasiska män, men dessa 18 personer ”förmodas ha afrikansk mtDNA, vilket också indikeras genom mönstret av mutationer av mtDNA-fragment”);
Asien(Kina, Vietnam, Laos, Filippinerna, Indonesien, Polynesien/Tonga) – 34 personer;
kaukasier(Europa, Nordafrika, Mellanöstern) – 46 personer;
australiensiska aboriginaler– 21 personer;
Nya Guinea– 26 personer.
Allt mtDNA delades upp i fragment med användning av restriktionsenzymer, vilket resulterade i totalt 467 oberoende mtDNA-sektioner, varav 195 hade skillnader i minst en person av alla 147. Med andra ord identifierades 195 polymorfa mtDNA-sektioner. I genomsnitt utfördes analys på 9 % av allt mtDNA. I allmänhet, för den tiden, för 25 år sedan, var detta ett ganska tekniskt avancerat arbete.
Därefter utförde vi parvisa jämförelser av de resulterande DNA-fragmenten mellan alla 147 deltagare, och fann att dessa parvisa skillnader varierade från noll till 1,3 mutationer per 100 nukleotider (0 till 1,3% skillnader), med ett totalt genomsnitt på 0,32% skillnader. Men det var nödvändigt att visa att dessa skillnader är störst bland afrikaner, så alla fem populationer delades in i kluster baserat på grupperna av parvisa skillnader i varje population. Det visade sig att
46 europeiska mtDNA:n divergerar i 36 kluster,
34 asiatiska mtDNA:n divergerar i 27 kluster,
21 australiska mtDNA:n divergerar i 15 kluster,
26 mtDNA från Nya Guinea divergerar i 7 kluster,
och 20 afrikanska mtDNA identifierades i ett kluster, vilket beslutade att sedan mänskligheten kom ut ur Afrika, borde det bara finnas ett kluster där. Detta är vad de skrev ner i anteckningen till tabellen i artikeln, där alla har många kluster, men afrikaner har bara ett.
Afrika: 0,36 %
Asien: 0,21 %
Australien: 0,17 %
Nya Guinea: 0,11 %
Europa: 0,09 %
Därefter översatte författarna dessa "mångfald" till kronologiska indikatorer, nämligen till åren då dessa territorier först beboddes. För att göra detta tog vi följande siffror för kalibrering: Australiens bosättning inträffade för 40 tusen år sedan, bosättningen i Nya Guinea för 30 tusen år sedan, bosättningen i Amerika för 12 tusen år sedan, och fann att mutationer i mtDNA inträffade vid en genomsnittlig hastighet på 2-4% (det vill säga 2-4 mutationer för varje 100:e nukleotider) per miljon år. Härifrån beräknade författarna till artikeln den genomsnittliga "åldern" för kluster i befolkningen:
Afrika: 90-180 tusen år
Asien: 53-105
Australien: 43-85
Nya Guinea: 28-55
Europa: 23-45
De gjorde det klumpigt, men siffrorna visade sig vara ganska rimliga (inom 100 % fel). Som efterföljande studier av andra författare, inklusive mina, som kommer att diskuteras nedan, visade, började afrikanska DNA-linjer för cirka 160 tusen år sedan, plus flera arkaiska afrikanska linjer (haplogrupper A0 och A00) med åldern cirka 180 respektive 210 tusen år; Asiatiska och europeiska härstamningar - från 64 tusen år sedan, Australien - från cirka 45-50 tusen år sedan, och de äldsta benresterna av moderna människor i Europa går tillbaka till 45 tusen år sedan (Benazzi et al, 2011; Higham et al. , 2011). Det är tydligt att författarna genomförde beräkningar med en noggrannhet på plus eller minus 100 %, men ändå fångas helhetsbilden relativt korrekt.
På liknande sätt beräknade författarna att den gemensamma förfadern till allt mtDNA levde för 143-285 tusen år sedan, och eftersom den gemensamma förfadern till allt afrikanskt mtDNA levde, enligt deras beräkningar, för 90-180 tusen år sedan, det vill säga äldsta av alla (även om den överlappar i ålder inom gränserna för räknefel), därför var det då han lämnade Afrika.
Märker du en förändring i begreppen? Författarna beräknar att människor utanför Afrika härstammar från en nyare gemensam förfader och postulerar att de därför kom ut ur Afrika. Som ett resultat avslutar författarna och detsamma är skrivet i abstraktet, en kvinna, den gemensamma förfadern till allt mtDNA på planeten, som "postulerat" (!), levde för 200 tusen år sedan (detta är redan en omvandling av 143 -285 tusen år sedan), och "förmodligen" (!) bodde hon i Afrika.
Allt började med den här artikeln. Jag upprepar, jag har ingen aning om hur en sådan artikel skulle kunna passera recensenterna och publiceras i tidskriften Nature, med dessa "postulerade" och "förmodligen", och i avsaknad av några uppgifter om framväxten av den moderna mänskligheten från Afrika, men det är precis så den här artikeln och började uppfattas både av media och i befolkningsgenetik, och därifrån i vetenskapen och bland den genomsnittliga människan - vilket betyder att det är ovedersägligt bevisat att den moderna människan kom ut ur Afrika. Det fanns praktiskt taget inga andra genetiska bevis, och varför? Allt har redan bevisats, eller hur?
Grundläggande misstag av kreatörer och supportrar
begreppet "mänskligheten lämnar Afrika"
Det finns ett grundläggande misstag som ständigt plågar befolkningsgenetiker. Om en population är mer genetiskt "diversifierad" än en annan, det vill säga kollektivt äldre, tenderar de att tro att den är förfäder till den andra. Men detta stämmer inte alls. Här måste vi titta på helheten av faktorer, och inte dra till en slutsats. Till exempel är en äldre bror "mer diversifierad" än en yngre, men det betyder inte att den yngre är en ättling till den äldre. De har bara en gemensam förfader, sin far. Detsamma gäller olika genealogiska konstruktioner, och om vi börjar jämföra syskonbarn och flyttar deras gemensamma förfäder till en gemensam farfar, farfarsfar, farfars farfar och så vidare, kommer vi att se att grenar av ättlingar kan avvika från gemensamt släktträd vid olika tidpunkter, men inte För att jämföra dem "efter ålder" linjärt, direkt med varandra, måste du säkert veta när deras gemensamma förfader levde.
Detta är tydligt om man tittar på ett vanligt träd. En tjock gren och en ung gren sitter i närheten, men det är inte alls nödvändigt att den unga kommer ut ur den gamla. Ofta är de oberoende upp till stammen; deras gemensamma förfader är en stam eller gren av ännu större tjocklek. Begreppet gemensam förfader behandlades inte alls i 1987 års tidning. Ett typiskt misstag för popgenetiker är "det jag ser är vad jag sjunger." Om de bor i Afrika nu betyder det att de alltid har bott där. Att den gemensamma förfadern till afrikaner och icke-afrikaner kunde ha levt utanför Afrika och migrerat dit i forna tider är inte ens övervägande av dem.
Det finns en annan grundläggande brist i popgenetikens synsätt baserat på att jämföra "mångfald". Mångfald är informativt i denna mening, som termodynamiken säger, endast i slutna system. New York är betydligt mer "diverserat" än till exempel Boston, men betyder det att Boston, en ättling till New York, kom ur det? Moskva är "mer mångfald" än Novgorod, men är Novgorod en ättling till Moskva? Inte alls. Raka motsatsen. Mångfald kommer ofta från att blanda olika populationer, eftersom systemet är öppet. Här i New York och Moskva finns en blandning, och mycket "mångfald" har samlats. Afrika är också ett öppet system. Många migrationer av olika haplogrupper flyttade dit i antiken och på relativt senare tid, och det är därifrån "mångfalden" kommer. Även haplogruppen R1b i sin del avancerade för cirka 5 tusen år sedan, nu bor de i Kamerun och Tchad (Cruciani et al, 2010), svarta, eftersom de blandas med lokala skönheter. Men haplogruppen fanns kvar, R1b. Har de bidragit till afrikansk "mångfald"? Naturligtvis, som många liknande migrationer till Afrika. Då och då dyker det upp akademiska artiklar som beskriver "inträdet i Afrika." Den senaste artikeln finns i samma tidskrift Nature i augusti 2013 (Hayden, 2013), som beskriver nyligen identifierade migrationer av populationer till Afrika för 3000 år sedan och för 900-1800 år sedan. Bidrog de till "mångfalden"? Otvivelaktigt. Dessutom gick de söder om Sahara, där författarna till artikeln från 1987 tog mtDNA-prover.
Atkinson gör samma misstag i en ny artikel (Atkinson, 2011), där han skriver: " genetisk och fenotypisk mångfald minskar med avståndet från Afrika... vilket stöder hypotesen om ett afrikanskt ursprung för människor" Låt oss titta på diagrammet som kommer att förklaras nedan. Till vänster är den afrikanska grenen, till höger den icke-afrikanska. Mångfalden (det vill säga antiken) minskar från vänster till höger, men inte alls eftersom den vänstra grenen är förfäders. De kommer båda från en gemensam förfader, som, som kommer att visas nedan, inte bodde i Afrika.
Ytterligare en illustration av nedgången i mångfald med avståndet från Afrika. Åldern för haplogrupp A i Afrika är cirka 160 tusen år, efter avvikelsen från alfa-haplogruppen. Åldern för haplogrupperna R1a och R1b på avstånd från Afrika är 20 tusen år respektive 16 tusen år, de bildades i Centralasien (Klyosov och Rozhanskii, 2012a; Klyosov, 2012). Mångfalden faller från Afrika till Centralasien? Faller. Är det för att R1a och R1b härstammar från afrikansk haplogrupp A? Absolut inte. Dessa är orelaterade händelser och system.
En analogi - om det finns ett vårdhem i en del av staden, är "mångfalden" där den högsta. Betyder det att alla i stan, inklusive dagiset tvärs över gatan, kom från äldreboendet? Inte alls nödvändigt. Dessa är orelaterade händelser och system. Detta kan vara sant om systemet var stängt, vilket betyder att ingen hade kommit in i staden på hundratals år. Men i verkligheten flyttar tusentals människor in till staden som inte har något att göra med de på äldreboendet och de på dagis, och till de förra hämtas även äldre från hela landet och från utlandet. Även om man mäter det är dess mångfald högst. Men det finns inga anor.
Här är exempel på "linjärt tänkande" angående "mångfald":
...Haplotypdiversiteten är högst i Afrika (Hellenthal et al., 2008);
Afrika har inte bara den högsta nivån av genetisk variation i världen, utan har också en betydande mångfald i språk, kulturer och miljöer (Campbell och Tishkoff, 2010).
Allt detta är sant, men det har ingenting att göra med huvuduppsatsen om mänsklighetens ursprung som förmodas vara i Afrika. Det här är bara ett par exempel av hundratals av samma sort.
För att diskutera "genetisk mångfald" måste vi känna till populationernas tillkomst, deras historia, och inte bara formellt mäta denna "mångfald" i olika populationer och "linjärt" jämföra den. Detta, jag upprepar, är befolkningsgenetikernas gissel. Varför är detta? Svag vetenskaplig skola, det finns ingen annan förklaring.

Diagram över utvecklingen av haplogrupper av modern mänsklighet. På den horisontella axeln finns huvudhaplogrupperna för mänsklighetens Y-kromosom, på den vertikala axeln är den absoluta tidsskalan. Den gemensamma förfadern till alfa-haplogruppen levde för cirka 160 tusen år sedan, den gemensamma förfadern till beta-haplogruppen (eller haplogrupperna B till T) levde för 64±6 tusen år sedan (från artikeln av A.A. Klyosov och I.L. Rozhanskii, Advances in Anthropology, 2012b). Diagrammet visar inte de arkaiska afrikanska linjerna A00 och A0 (den senare har nu ersatt A1b-linjen i nomenklaturen i diagrammet till vänster), det uppdaterade haplogruppsträdet kommer att visas nedan.
Det finns (åtminstone) ytterligare en anledning till den lägre "mångfalden" bland icke-afrikaner. För cirka 64 tusen år sedan passerade deras förfäder "befolkningsflaskhalsen". Med andra ord, som ett resultat av någon katastrof, dog nästan alla icke-afrikaner eller degenererade, och endast en liten grupp av dem överlevde. Bildligt talat överlevde i slutändan bara ett pars ättlingar, och nu konvergerar alla genealogiska linjer för nästan alla män på planeten till dem. Vilken typ av katastrof eller annan olycka, som en pest, som hände är okänt, och två hypoteser har störst tyngd - utbrottet av Toba-vulkanen, den största kända i mänsklighetens historia, för cirka 70 tusen år sedan, och ett köldknäpp i norra halvklotet. Klimatologer säger att den katastrofala karaktären av köldknäppet är högre än Toba-utbrottet. Hur som helst, här är vad som hände (se diagram nedan).
Det är tydligt att om vi mäter "mångfalden" av grenarna till vänster (afrikanska baserat på dess representanters nuvarande bostad) och till höger (icke-afrikanska, också baserat på den nuvarande bostaden), så kommer den första att vara äldre. Men den högra grenen kommer inte ut från den vänstra, de har en gemensam förfader, alfa-haplogruppen. Som du kan se förklarar diagrammet alla resultat från 1987 års Cannes-tidning, men utan att lämna Afrika. Ytterligare bevis på riktigheten av detta diagram kommer att ges nedan.
Multiplikation av fantasier när de sprids
begreppet "mänskligheten lämnar Afrika"
Med tiden dök figuren på något sätt upp ur tomma luften att den moderna människan kom ut ur Afrika för 70 tusen år sedan, och den började också citeras i hundratals akademiska artiklar, så att man förlorade slutsatserna om vem som sa det först och när . Och vem brydde sig? Kom den moderna människan ut från Afrika? Den kom ut för länge sedan och visas obestridligt redan 1987. När kom den ut? Så, alla skriver att för 70 tusen år sedan har det också visats för länge sedan och ovedersägligt. Några andra frågor? Vem tror inte på konsensus? Låt oss ta en titt på den här och sedan vidta åtgärder.
Och dussintals och hundratals akademiska artiklar dök upp, den första frasen där vanligtvis var " Som bekant dök den anatomiskt moderna människan upp från Afrika för cirka 70 tusen år sedan." Men denna datering var också "flytande", och nedan finns exempel på olika dateringar av "utträdet från Afrika" i olika artiklar. En liten hemlighet – ingen av dem var faktiskt uträknade. Alla är bara ur det blå. Ja, och tills nyligen fanns det ingen motsvarande beräkningsapparat, men den som fanns - läsaren har redan sett hur det är och vilken noggrannhet det är.
50 tusen år sedan (Jobling & Tyler-Smith, 2003);
50 tusen år sedan (Thomson et al, 2000);
50-60 tusen år sedan (Shi et al., 2010);
50-60 tusen år sedan (Mellars, 2011);
50-70 tusen år sedan (Hudjasov et al., 2007);
50-70 tusen år sedan (Stoneking & Delfin, 2010);
60 tusen år sedan (Li & Durbin, 2011);
60 tusen år sedan (Henn et al., 2011);
60 tusen år sedan (Wei et al., 2013);
60-70 tusen år sedan (Ottoni et al., 2010);
60-80 tusen år sedan (Forster, 2004);
54±8 tusen år sedan (Forster et al., 2001);
60 tusen år sedan (Stewart & Stringer, 2012);
45-50 tusen år sedan (Fernandes et al., 2012);
50-65 tusen år sedan (Behar et al., 2008);
50-60 tusen år sedan (Cann, 2013);
60 tusen år sedan (Chiaroni et al., 2009);
50-75 tusen år sedan (Patin et al., 2009);
50 tusen år sedan (Edmonds et al., 2004);
45 tusen år sedan (Moorjani et al., 2011);
50-70 tusen år sedan (Xue et al., 2005);
70-80 tusen år sedan (Majumder, 2010);
40 tusen år sedan (Campbell och Tishkoff, 2010);
50 tusen år sedan (Poznik et al, 2013);
55-70 tusen år sedan (Soares et al., 2009);
mellan 40 och 70 tusen år sedan (Sahoo et al., 2006);
mellan 35 och 89 tusen år sedan (Underhill et al., 2000);
mellan 80 och 50 tusen år sedan (Yotova et al., 2011);
mellan 50 och 100 tusen år sedan (Hublin, 2011);
mellan 27-53 och 58-112 tusen år sedan (Carrigan och Hammer, 2006);
70-60 tusen år sedan (Curnoe et al., 2012);
~110 tusen år sedan (Francalacci et al, 2013);
200 tusen år sedan (Hayden, 2013).
Faktum är att inget datum kan rimligen anges. Hon finns helt enkelt inte. Och alla dessa dateringar ovan är till ingen nytta för någon, de ger ingenting och svarar i princip inte på några frågor. Det är fortfarande samma mantra.
Vad de experimentella uppgifterna verkligen säger
och deras bredare tolkning?
Låt oss stanna ett tag med kritik och se – vad finns det? Om moderna icke-afrikaner inte är ättlingar till forntida afrikaner, var följer då detta? Vems ättlingar är de?
Arkeologiska och paleontologiska data om Afrika kommer att behöva kasseras med beklagande. De är informativa av allmänna skäl, men vi vet inte om de påträffade benresterna hade några överlevande ättlingar. Kanske observerar vi resterna av avslutade linjer. Tills haplogrupperna och haplotyperna för dessa benrester är etablerade, kommer de inte att berätta något om kontinuiteten hos dessa evolutionära linjer. Vidare vet vi inte var dessa benrester kom därifrån. Kanske har deras nära förfäder migrerat till Afrika. Ja, om det var möjligt att lämna Afrika, så var det också möjligt att komma in där. Dessutom är många exempel på migrationer till Afrika kända. Många dateringar av forntida benrester är felaktiga, och exempel kommer att ges nedan. Många påstod sig vara "urgamla" Homo sapiens» har uttalade arkaiska drag, och deras klassificering som Homo sapiensär generellt kontroversiell eller helt enkelt felaktig. Många fynd hänför sig inte alls till benrester, utan till platser, grottor, snäckor som hittats där och stenredskap. Det är inte känt vem som var där alls, och ockran som hittats där säger inte heller något. Neandertalarna i Eurasien använde också stenredskap och ockra för sina egna syften.
Så frågan om hur forntida afrikaner och icke-afrikaner är släkt är mer lämpligt åtgärdade genom att titta på bådas DNA. Om dessa data också stöds av arkeologi-antropologi är detta underbart, men än så länge är sådana data få, om några. Låt oss titta på dem.
DNA-data kan ses på tre sätt, vilket i princip ska ge ömsesidigt överensstämmande data. Dessa är (1) haplotyper och haplogrupper av den humana Y-kromosomen, (2) humant mtDNA och (3) det mänskliga genomet. Det senare betyder egentligen en bild av irreversibla mutationer i DNA, som kan tolkas i relation till mänsklig evolution, som visar riktningen för flödet av mutationer och uppkomsten av nya under den evolutionära utvecklingens gång. Till exempel, i genomet hos både neandertalare och moderna människor finns det många av samma mutationer som också finns i schimpansernas DNA. Det betyder att dessa mutationer är från den gemensamma förfadern till människor och schimpanser. Men om vi också har mutationer från neandertalare som schimpanser inte har, så kan det betyda att neandertalarna är vår direkta förfader. Sådana mutationer identifieras antingen inte, eller så finns det väldigt få av dem och de är kontroversiella. Uppgifterna revideras för närvarande. De nyligen utropade 1-4% som förmodas i den moderna människan från neandertalaren revideras nu också. Troligtvis är de felaktiga.
På samma sätt delar både icke-afrikaner och afrikaner mutationer från en gemensam förfader med schimpanser. Det finns många av dem, och de är inte intressanta för att avgöra om vi härstammar från afrikaner. Dessa mutationer måste filtreras bort. Men har vi mutationer som afrikaner har, men inte schimpanser – det är en fråga som måste besvaras. Detta svar, om det erhålls, måste överensstämma med data om haplotyper och haplogrupper av Y-kromosomen och mtDNA. Så är studiet av frågan om vilka den moderna mänsklighetens evolutionära vägar bör struktureras.
I själva verket har detta svar redan mottagits - det finns inga "afrikanska" mutationer i oss som de förvärvat under de senaste 150-200 tusen åren. Det finns många mutationer från en gemensam förfader med schimpanser, som är miljontals år gamla, i vårt DNA, men det finns inga mutationer från afrikaner som förvärvats under de senaste 160 tusen åren i vårt DNA.
Detta är vad vi kommer att prata om.
Så vad säger experimentella data om detta? Låt oss börja med mänskliga haplotyper och haplogrupper som projicerades till tider för mer än 100 tusen år sedan. Projicerat - eftersom det inte finns några fossila haplotyper och haplogrupper från den tiden. Även om den ännu inte har identifierats, är uppgiften tekniskt sett mycket svår, eftersom mänskligt DNA under sådana tider sönderdelas nästan helt, särskilt under påverkan av mikroorganismer. DNA från en neandertalare (mer exakt, en neandertalare) för 45 tusen år sedan har isolerats och till stor del dechiffrerats, men DNA från en person för 160 tusen år sedan är en uppgift som är storleksordningar svårare.
Vad gör man i sådana fall? Haplotyper bestäms i moderna populationer av män om analysen utförs på Y-kromosomen. Men provet analyseras inte enligt någon "Nya Guinea" eller "afrikansk" befolkning, som kan vara helt heterogen, utan bland bärare av en viss undergrupp av en haplogrupp, det vill säga bland människor förenade av en specifik uppsättning mutationer. De är släktingar, och för dem är det ganska exakt beräknat när deras gemensamma förfader levde. Till exempel, bland moderna afrikaner finns det en ganska representativ grupp människor som, enligt klassificering, faller i subclade A1b1b2b, markerad med färg (tredje från botten) på trädet av subclades av haplogrupp A nedan. Det här trädet visar hierarkin av underkläder av haplogrupp A, det vill säga utvecklingen av haplogrupp A. Du kan se hur trädet grenar - den äldsta haplogruppen A00 har flyttat bort från stammen, dess grenar (underkläder) är fortfarande okända. Stammen fortsätts av haplogrupp A0-T, som delas upp i två undergrupper - A0 och A1; Al divergerade i sin tur in i Ala och Alb; A1b – till A1b1 och VT. Den kombinerade haplogruppen BT, som kommer att visas senare, är mycket avlägsen från haplogrupperna i "A"-serien, och även i "A"-serien är det inte klart vilka undergrupper som kan kallas afrikanskt ursprung. Hittills verkar det bara haplogrupperna A00 och A0, det vill säga den första och tredje från toppen (flyttar sig bort från stamgrenen som leder från en gemensam förfader med schimpanser för miljoner år sedan, till den icke-afrikanska haplogruppen BT) på trädet som visas nedan, och de senares underkläder (A0a, A0b, A0a1, A0a2, A0a1a och A01ab) kan anses vara afrikanskt ursprung eller ankomst till Afrika för över 100 tusen år sedan. Resten, som börjar med A1 (som ligger på samma stamgren), delar sig i afrikanska (förgrenar sig åt sidan) och förmodligen icke-afrikanska (stam) grenar.

Låt oss återigen, tydligare, titta på hur trädet av haplogrupper förgrenar sig, hur varje gren divergerar till gafflar och hur en del av gaffeln lämnar (migrerar) till Afrika, medan den andra delen förblir utanför Afrika, och återigen divergerar in i nästa. gaffel. Med andra ord kom migrationerna till Afrika i vågor. Som ett resultat kan en icke-afrikansk stam spåras, vilket leder till dig och mig, läsare, och från vilken afrikanska skott sträcker sig åt sidan. Vi kom inte från dessa rymningar. Här bör det noteras att termerna "stam", "stam" och "förgrening åt sidan" väljs villkorligt, och man kan tvärtom kalla afrikanska grenar för stam och icke-afrikanska - förgrenade åt sidan. Dessa begrepp är faktiskt symmetriska.
Gaffel 1- från den huvudsakliga evolutionära Y-kromosomala "stammen", som kommer från gemensamma förfäder med primater (schimpanser, gorilla, orangutang, makaker) för cirka 300-600 tusen år sedan, avgår en gren av neandertalarna ( Homo neanderthalensis); de var inte afrikaner, i alla fall, inga spår av dem hittades i Afrika, så vi kan anta att den vanliga stammen för 300-600 tusen år sedan var ett icke-afrikanskt släkte Homo.
Gaffel 2– den äldsta av de hittills upptäckta grenarna av buketten av haplogrupp A, haplogrupp A00, avgår från stammen för cirka 210 tusen år sedan (nu lever alla dess upptäckta bärare i Afrika som en del av Mbo-stammen, eller är afroamerikaner ; ingen information om deras antropologi eller anatomi har hittats lyckades; i artikeln som ger deras haplotyper, inte ett ord om detta).
Gaffel 3– stammen når haplogrupp A0-T (förmodligen icke-afrikansk), som divergerade för cirka 180 tusen år sedan in i den afrikanska haplogruppen A0 och förmodligen icke-afrikansk A1; med andra ord, en annan afrikansk haplogrupp A0 förgrenar sig från stammen.
Plugg 4– Icke-afrikansk haplogrupp A1 divergerar till afrikansk A1a och förmodligen icke-afrikansk A1b; med andra ord, en annan afrikansk haplogrupp A1a avgår från stammen.
Gaffel 5– Icke-afrikansk haplogrupp A1b divergerar i afrikansk A1b1 och icke-afrikansk BT (beta-haplogrupp i det första diagrammet); med andra ord, en annan afrikansk haplogrupp A1b1 avgår från stammen.
Nu - en mycket viktig punkt i vår övervägande. Gafflarna 3, 4 och 5 avviker från haplogrupperna A0-T, A1 respektive A1b.
Från den första sträcker sig A0 (som hittades i Afrika) och A1 (vars bärare ännu inte har hittats någonstans) åt sidan. Vi icke-afrikaner härstammar från A1 (och härstammar inte från A0; vi har inte dess mutationer).
På väg bort från A1 är A1a (som finns i Afrika) och A1b (vars bärare ännu inte har hittats någonstans). Vi icke-afrikaner härstammar från henne. I vår Y-kromosom finns mutationer från A1b, men inte från A1a.
Avgrenade från A1b är A1b1 (som finns i Afrika, Europa och Asien) och BT, från vilka alla icke-afrikanska haplogrupper uppstod, inklusive de viktigaste europeiska haplogrupperna R1a, R1b, I1, I2, N1c1.
För att "bevisa" att alla människor på jorden kom ut ur Afrika (i form av sina förfäder, naturligtvis), förklarar anhängare av konceptet "ut ur Afrika" alla dessa tre nodhaplogrupper - A0-T, A1 och A1b "afrikansk". Jag upprepar att ingen av dem har hittats i Afrika. Men detta stör inte "supportrarna". Läsaren har redan insett att det finns andra tekniker som inte kan kallas vetenskapliga. De är förklarade afrikanska, och "supportrarna" säger - ja, se, alla europeiska och asiatiska haplogrupper kommer från afrikanska, från A0-T, A1 och A1b. Det är det, konceptet att "gå ur Afrika" har bevisats.
I själva verket är detta inget bevis, utan ett hån mot vetenskaplig övervägande och sunt förnuft. Det är mycket mer troligt att dessa tre haplogrupper inte alls är afrikanska, och att deras bärare levde utanför Afrika. Då är sambandet mellan den ljushyade neandertalarens förfader (mer om detta nedan) och de ljushyade moderna människorna lätt att förklara. Det är lätt att förklara avfärden till Afrika - efter gaffelavvikelserna - för bärarna av haplogrupperna A0, A1a, A1b1, som nu till övervägande del lever i Afrika. De enorma tidsavstånden mellan afrikanska och icke-afrikanska haplogrupper är lätta att förklara, eftersom de konvergerar till avlägsna gemensamma förfäder och inte kommer direkt från varandra (då skulle avstånden vara ungefär 60-70 tusen år, men de är faktiskt 250- 300 tusen år. I princip kan icke-afrikanska härstamningar uppstå från afrikanska så att de är åtskilda med 250-300 tusen år. Och anhängarna av "utträdet från Afrika" själva proklamerar hela tiden att utträdet ägde rum 60-70 tusen år år sedan.De visste inte, och vet inte, att där är avståndet faktiskt 4-5 gånger större.
Därför, i beskrivningen av gafflarna ovan, skriver jag överallt "förmodligen icke-afrikansk haplogrupp" A0-T, A1, A1b.
Varhelst neandertalarnas förfäder och de som de divergerade med under evolutionen bodde (det vill säga de som fortsatte "huvudstammen" av Y-kromosomens evolutionära träd), migrerade bärare av haplogrupperna A00, A0, A1a, A1b1 från dem till Afrika och fortsatte sin utveckling där, tog emot många senare migranter till Afrika och ökade därmed den afrikanska "mångfalden".
I allmänhet kan fyra stora forntida migrationer till Afrika räknas under de senaste flera hundra tusen åren - haplogrupp A00 för ungefär 210 tusen år sedan, haplogrupp A0 för ungefär 180 tusen år sedan, haplogrupp A1a för ungefär 160 tusen år sedan, haplogrupp A1b1 för ungefär 70 tusen år sedan sedan . Naturligtvis förekom senare migrationer, till exempel för 3000 och 900-1800 år sedan, beskrivna i (Hayden, 2013), som också ökade den "genetiska mångfalden" i Afrika, så "mångfald" är inte ett argument för "fäders hemland" .
Jag har redan nämnt att bärare av haplogrupp A1b1 lever i Afrika, Europa och Asien. Tydligen är det därför som A1b1b2b-M13-subkladden i Haplogroup A-projektet visar sig vara den mest talrika. Det divergerar i två huvudgrenar - arabiska och europeiska. Vi vet inte vem som var förfadern till dessa grenar och var han bodde, men grenen är ganska ytlig, det vill säga att den passerade befolkningsflaskhalsen relativt nyligen. Dess haplotyper representerar en värdefull informationskälla, eftersom de placerar även en ytlig (i tiden) gren i området för alla haplotyper av mänskligheten. Haplotyper och efter flaskhalsen kunde populationer inte uppstå spontant, de kunde bara fortsätta evolutionen från de äldsta gemensamma förfäderna. Analys av haplotyper med användning av de långsammaste, mest stabila 22 markörerna av Y-kromosomen (Klyosov, 2011) visar att den gemensamma förfadern till den arabiska grenen av A1b1b2b-subkladen hade haplotypen
12 11 11 9 11 10 10 9 12 12 7 12 8 0 13 11 16 9 14 9 11 11
Och den europeiska grenens gemensamma förfader hade haplotypen
12 11 11 9 11 10 10 9 12 12 7 10 8 0 13 11 16 10 14 9 11 11
Det finns bara tre mutationer mellan dem, vilket placerar den gemensamma förfadern till de arabiska och europeiska linjerna för cirka 7 170 år sedan, med en felmarginal på plus eller minus fem procent. För syftet med vår beskrivning är dessa beräkningar ännu inte särskilt viktiga, eftersom det är tydligt att ovanstående haplotyper ligger nära varandra.
Låt oss jämföra dessa haplotyper med den förfäders afrikanska haplotypen av grupp A00:
13 11 12 10 11 16 10 9 14 14 8 8 8 9 12 11 12 8 12 12 11 11
Denna jämförelse visar en skillnad på redan 30 och 29 mutationer, det vill säga den separerar de gemensamma förfäderna för dessa haplotyper med minst 286-308 tusen år (beräkningsformler publiceras i arbetet), och placerar den gemensamma förfadern till haplogrupp A00 vid för cirka 210 tusen år sedan. Sambandet mellan antalet mutationer och tid är inte linjärt, utan maktlag, eftersom vissa mutationer under långa tider återkommer, och en lämplig statistisk korrigering införs för detta i beräkningarna (Klyosov, 2009; Klyosov, 2012). Haplotyper av haplogrupp A00 erhölls från den svarta Mbo-stammen som bor i Kamerun och från en afroamerikan, antagligen hämtad för århundraden sedan från samma stam (Mendez et al, 2013).
Om vi nu jämför dessa haplotyper med den förfäders haplotypen av haplogrupp B
11 12 11 11 11 10 11 8 16 16 8 10 8 12 10 11 15 8 12 11 12 11
Sedan kommer vi att se 29 mutationer från haplogrupp A00, och nästan samma antal - 29 och 27 mutationer - från de arabiska och europeiska grenarna av haplogrupp A1b1b2b. Detta är minst 286-248 år mellan de gemensamma förfäderna till haplogrupp A och B. Denna kolossala separation i tiden tillåter inte haplogrupp B att vara en ättling till haplogrupp A. Men att ha en gemensam förfader för 160 tusen år sedan och avvika från den genom att 250-300 tusen år - Kan. Detta överensstämmer återigen med diagrammet ovan. Dessa haplogrupper kan inte jämföras "linjärt" bara för att de är visuellt nära på haplotypträdet, precis som det är omöjligt att jämföra grenarna på ett träd i en skog "linjärt", med avståndet mellan dem, bara för att de råkar vara närliggande. Och i närheten kan det finnas grenar av björk och gran som växer i grannskapet.
Så, haplogrupp B är mycket avlägsen från haplogrupp A, genom 27-29-30 mutationer. Men det är inte så långt borta från de europeiska (till stor del) haplogrupperna R1a och R1b, med endast 12 respektive 10 mutationer:
11 12 13 11 11 12 11 9 15 16 8 10 8 12 10 12 12 8 12 11 11 12 (R1b-M269)
12 12 11 11 11 11 11 8 17 17 8 10 8 12 10 12 12 8 12 11 11 12 (R1a-Z280)
Dessa haplotyper själva (R1b och R1a) är åtskilda av endast 8 mutationer, vilket motsvarar livslängden för deras gemensamma förfader (haplogrupp R1) för ungefär 26 tusen år sedan. Den gemensamma förfadern till haplogrupp B levde för cirka 50 tusen år sedan, och den bildades inte från haplogrupp A, de är oberoende DNA-genealogiska linjer som kommer från en gemensam förfader - alfa-haplogruppen, för 160 tusen år sedan.
Det finns andra bärare av haplogrupp A i Europa, även om få har hittats hittills. För några år sedan fanns det en akademisk artikel med titeln "Afrikaner i Yorkshire?" (King et al, 2007), som beskriver en familj av haplogrupp A-bärare i England som inte hade någon aning om att de hade afrikanska anor i manlig linje. Deras grundläggande 17-markörshaplotyp visade sig vara följande (i markörordning DYS393, 390, 19, 391, 388, 439, 389-1, 392, 389-2, 437, 438, 434, 435, 436, 436 , 461, 462):
14 23 17 10 10 11 12 11 17 14 8 12 12 11 11 12 12
Och den arabiska grenen av subclade A1b1b2b som beskrivs ovan
13 21 15 9 11 12 13 11 18 16 10 9 11 11 11 13 13
Mellan dem finns 20 mutationer på 17 markörer, vilket motsvarar minst 19 tusen år före deras gemensamma förfader, de engelska och arabiska haplotyperna av haplogrupp A. Det är omöjligt att säga vem som i detta fall flyttade vart - antingen till Afrika eller från Afrika . Det kan finnas vilka scenarier som helst. En anhängare av konceptet "ut ur Afrika" kommer omedelbart att säga att de kom ut ur Afrika. Detta är den psykologiska attityden.
Debatten under de senaste två åren angående diagrammet ovan
När artikeln som innehåller diagrammet ovan och dess tolkning publicerades i maj 2012 i tidskriften Advances in Anthropology möttes den till en början av motstånd från populationsgenetiker. Specifikt har tre huvudfynd varit kontroversiella: (1) Afrikanska och icke-afrikanska DNA-linjer skilde sig åt för ungefär 160 tusen år sedan, och det finns ett motsvarande betydande avstånd mellan dem; (2) icke-afrikanska DNA-linjer är inte ättlingar till afrikanska haplogrupper A00, A0, A med undergrupper; och, som en konsekvens, (3) den moderna mänskligheten inte hade någon "utträde från Afrika", åtminstone under de senaste 200 tusen åren. Och om det fanns, var det fram och tillbaka, motverkar migrationer, och nästa "fram och tillbaka" migration gav inte upphov till modern mänsklighet. I vilket fall som helst är de i detta avseende likvärdiga.
Det måste sägas att detta avslag inte alls kom till uttryck i den akademiska vetenskapliga pressen. Befolkningsgenetiker är fulla av vatten. Passionerna var höga i engelskspråkiga forum och i informella diskussioner. Det tillkännagavs att detta diagram och följaktligen dess slutsatser helt motsäger konsensus om mänsklighetens utträde från Afrika, och motsäger alla publicerade diagram och haplogruppträd som erhållits genom genomisk forskning. Det tillkännagavs också att icke-afrikanska härstamningar kommer från haplogrupper med indexet "A", vilket betyder afrikansk. Detta förklarades vara oförenligt med utvecklingen av kvinnligt mtDNA, där (nu) icke-afrikanska linjer också dök upp från Afrika, och manliga och kvinnliga haplogrupper måste ha dykt upp från Afrika tillsammans.
I själva verket var alla dessa invändningar i princip felaktiga. Motståndarna antingen inte ville eller kunde inte lista ut det och försökte som vanligt "ta det i halsen." Låt oss ta en titt och se till att det faktiskt inte finns några motsägelser, och motståndarna upprepade helt enkelt memorerade mantran, vilket alltför ofta accepteras inom populationsgenetik.
Den senaste boken "Evolutionary Genetics of Humans" -
korrekta uppgifter, felaktig tolkning
Vi öppnar en ny bok - "Human Evolutionary Genetics", författarna Jobling, Hollox, Hurles, Kivisild, Tyler-Smith, publicerad 2014 (det stämmer, förlaget har hoppat framåt sex månader), kapitel 9 - "The Origin of Modern Man” , s. 304-305. Avsnitt "Mitokondriellt DNA". Citat: "Forskning har visat slående egenskaper: fullständig separation av afrikanska och icke-afrikanska härstamningar." Avsnitt "Y-kromosomer". Citat: "Även om det är mindre detaljerat än mtDNA, har studier visat nära paralleller: fullständig separation av afrikanska och icke-afrikanska linjer."
Som vi kan se finns det inga motsägelser med diagrammet ovan. Men författarna blåser redan upp sina tolkningar, baserat på data från 2000 – både på mtDNA och på Y-kromosomen. Således anses Y-kromosomal haplogrupp B vara afrikansk, och det skrivs att motsvarande gren innehåller "både afrikanska och icke-afrikanska DNA-linjer." Vi tittar på diagrammet - ja, haplogrupp B är i samma kluster med icke-afrikanska haplogrupper, och vi visade ovan att den är avlägsen från afrikanska haplogrupper, och är i samma kluster med icke-afrikanska, med en gemensam förfader. Varför kallade författarna det "afrikanskt"? Ja, många bärare av haplogrupp B Nu bor i Afrika. Kommer du ihåg hur jag skrev om populationsgenetiker? "Det jag ser är vad jag sjunger." De har både haplogrupp B och icke-afrikanska linjer i ett kluster, vilket betyder "utgång från Afrika." Och de är alla där, i det klustret, icke-afrikanska. Ja, även om det fanns en icke-afrikansk linje tillsammans med icke-afrikanska, varför är det nödvändigt att "lämna ut ur Afrika"? Varför inte lika väl "inträde i Afrika"? Och så, de marscherar längs den slitna stigen, svaret är känt i förväg. Författarna till boken anger dateringen av detta kluster, som innehåller icke-afrikanska haplogrupper och haplogrupp B (även icke-afrikanskt ursprung), för 52 ± 28 tusen år sedan. I min artikel – för 64±6 tusen år sedan. Var finns motsättningen?
Samma författare ger dateringen av alla DNA-linjer - 172 ± 50 tusen år sedan. Ja, i min artikel för 160±12 tusen år sedan. Var finns motsättningen? Det vill säga, befolkningsgenetiker tvistar inte i huvudsak, inte med data i handen, utan helt enkelt "i princip", för avvisningens skull. Det vanliga.
När det gäller mtDNA ger författarna symmetriskt samma tolkningar som med Y-kromosomen - en liknande gren som innehåller "afrikansk mtDNA" (eftersom de bor där nu) och icke-afrikansk - vilket betyder "utträde från Afrika", och dateringen av denna "blandade" gren är mellan 31 och 79 tusen år sedan, med en median på 40 tusen år sedan, dateringen av hela mtDNA är mellan För 40 och 140 tusen år sedan är medianen 59 tusen år sedan. Författarna diskuterar inte skillnaden i datering mellan Y-kromosomala data och mtDNA, men varför? Slutsatsen har länge varit klar - "mänsklighetens utträde från Afrika." Samma slutsats, om än i en försiktig form, finns också i kapitlets avslutning. Den talar också om "högre genetisk mångfald i Afrika", och det faktum att människor dök upp i Afrika för ungefär 200 tusen år sedan och utanför Afrika - efter 45 tusen år sedan. Här pratar vi också om experternas samförstånd om att "gå ur Afrika". Vi har sett att alla dessa (eller liknande) datering och "diversiteter" förklaras av diagrammet ovan, men popgenetiker vill inte ha andra förklaringar. De har en "konsensus".
Fortsatt berättelse från Cannes (1987)
"om att lämna Afrika", men utan Cannes (1991)
Fortsättningen på artikeln av Cann och andra (Cann, Stoneking och Wilson, 1987), som vi diskuterade ovan, är intressant på sitt sätt. En ny artikel kom fyra år senare (Vigilant et al, 1991), med Cannes inte längre bland författarna, utan två tidigare medförfattare, Stoneking och Wilson, med tre nya författare. Artikeln från 1991 rapporterar att Kann et al.s (1987) uppsats motarbetades starkt av många experter på grund av det faktum att mänsklighetens gemensamma förfader förmodligen bodde i Afrika, och medger att Kann et al.s (1987) artikel hade många svaga länkar. Författarna (varav två var författarna till det svaga, som de medger, arbete) listar dessa svaga länkar genom hela stycket - det finns en indirekt metod för mtDNA-jämförelse, och ett litet urval, huvudsakligen bestående av amerikaner av afrikansk härkomst, och en medvetet olämplig metod "mittpunkt" som tillämpas av författarna till artikeln från 1987, och bristen på statistisk bearbetning av erhållna data och "otillräcklig kalibrering" av mutationshastigheten i mtDNA och andra. Med andra ord, denna uppenbart svaga artikel, som författarna själva medgett, utgjorde grunden för teorin om "exit from Africa". Men processen hade redan börjat, så den efterföljande artikeln (1991) hade som mål att fortfarande rättfärdiga konceptet "ut ur Afrika", att icke-afrikaner härstammade från afrikaner, och i själva verket ersätta den svaga, kritiserade artikeln från 1987.
Och vad var den motiveringen? Visa att afrikansk mtDNA är äldre än icke-afrikansk mtDNA. Men detta är återigen en fortsättning på det eviga fundamentala misstaget hos befolkningsgenetiker, att om en population är äldre än en annan, så antas den första populationen vara urfader i förhållande till den andra. Låt oss titta igen på samma diagram ovan - den vänstra grenen är äldre än den högra, men den är inte förfäder till höger. De har en gemensam förfader. Och detta grundläggande misstag av popgenetik har upprepats under de kommande 25 åren, fram till nu. Om och om igen upprepar författarna till artikeln (1991) att den afrikanska grenen är äldre än den icke-afrikanska grenen, vilket betyder att den är förfäder, utan att inse att detta inte alls är bevis på "anor". Min farbror är "äldre" än mig, men han är inte min förfader.
I uppsatsens slutsats (1991) skriver författarna: vi har presenterat de starkaste bevisen för att vår gemensamma förfader levde i Afrika för 200 tusen år sedan. Faktum är att, som läsaren för länge sedan har insett, vad dessa bevis egentligen handlade om var att den befintliga raden av människor som nu bor i Afrika är äldre än den befintliga linjen av människor som bor utanför Afrika. Dessa "bevis" säger ingenting om "anor". För att göra detta är det nödvändigt att jämföra populationernas haplotyper (vilket författarna till artikeln från 1991 inte gjorde, och popgenetiker fortfarande inte gör) och deras snipmutationer (vilket författarna inte gjorde), och det senare visar också att våra förfäder inte kom från Afrika. Detta diskuteras i nästa avsnitt.
SNP-mutationer visar att vi är det
inte ättlingar till afrikaner i haplogrupperna A eller B
Låt oss gå vidare till en färsk artikel (Scozzari et al, 2012), som ofta citeras som ett exemplariskt arbete om det afrikanska genomet och skälen till "mänsklighetens utträde från Afrika." Faktum är att artikeln tillkännager upptäckten av 22 nya irreversibla mutationer i den mänskliga Y-kromosomen, bekräftelsen av 146 kända mutationer och konstruktionen av ett nytt, förbättrat träd av haplogrupper och subklader av afrikaner med en övergång till den icke-afrikanska delen av träd, och specifikt den konsoliderade haplogruppen ST. Detta är hela högra sidan av trädet ovanför i diagrammet, från haplogrupp C till R2. Författarna till artikeln kallar det "att komma ur Afrika." Låt oss se om detta är sant. Träd av haplogrupper och undergrupper från artikeln (Scozzari et al, 2012):

Trädet av de äldsta haplogrupperna och subkladerna, som ges i artikeln (Scozzari et al, 2012). Antalet irreversibla mutationer (SNP, Single Nucleotide Polymorphism eller SNP) som definierar vissa subklader visas. Det kan ses att mer än hälften av subkladerna tillhör haplogrupp A, som författarna anser vara afrikanska. Alla andra undergrupper, utom en, tillhör haplogrupp B, som författarna också anser vara afrikanska. Haplogruppen nere till höger, CT, består, enligt författarna, av 19 DNA-linjer, alla icke-afrikanska. Bilden är klickbar.
Låt oss uppmärksamma några funktioner hos trädet i den här bilden. Det börjar (mer exakt, det fortsätter den evolutionära stammen av den mänskliga Y-kromosomen) i den övre vänstra delen av diagrammet, omedelbart finns den första divergensen, eller gaffeln (haplogrupp A0-T, även om detta namn inte visas på diagrammet ), till haplogrupp A1b (som i diagrammet) med subklader, å ena sidan, och på resten av trädet, å andra sidan. Med andra ord, den första afrikanska grenen avgår från trädet, och inga icke-afrikaner (haplogrupp ST) stiger ner från det. Artikeln använder den redan föråldrade nomenklaturen från 2011, och det som i artikeln betecknas som A1b heter nu A0, med SNP:er V148, V149 och andra visas på den översta raden i diagrammet (se även haplogrupp A-trädet ovan).
Vid nästa gaffel (haplogrupp A1) går de afrikanska haplogrupperna A1a och A1a1 med sina SNP:er M31, P82, V4 och andra åt sidan, och resten av trädet går till andra sidan. Icke-afrikaner (haplogrupp ST) härstammar inte heller från den andra afrikanska grenen (A1a med subclade).
Den tredje gaffeln är haplogrupp A1b enligt nuvarande klassificering. Afrikanska haplogrupper A2 och A3 med undergrupper (föråldrad nomenklatur) går bort från det, nu är det A1b1 med SNP V249/L419, som ytterligare divergerar i undergrupper A1b1a-V50 (tidigare A2) och A1b1b-M32 (tidigare A3), båda med undergrupper . Bland de senare är A1b1b2b-M13 subclade, samma som de arabiska och europeiska DNA-linjerna kommer från, vars haplotyper vi diskuterade ovan. Den andra grenen av denna gaffel från haplogruppen A1b är den sammansatta haplogruppen BT, som visas längst ner i figuren från artikeln (Scozzari et al, 2012). Det är tydligt att denna BT-haplogrupp inte på något sätt härrör från de "afrikanska" haplogrupperna med index A, som alla är överst i denna siffra. Ordet "afrikansk" här måste sättas inom citattecken, eftersom bland deras undergrupper finns samma europeiska och arabiska grenar, och den europeiska är huvudsakligen England, Irland, Skottland, Turkiet (även om endast 3% av Turkiet är geografiskt beläget i Europa), arabiska – främst Saudiarabien, och haplotyper från England, Schweiz, Finland och andra länder.
Naturligtvis kan man hävda att de europeiska och asiatiska haplotyperna av haplogrupp A en gång lämnade Afrika tillsammans med migranter därifrån, men det kan lika gärna hävdas att de kom till Afrika på samma sätt. Så dessa argument går inte igenom, även om endast "ut ur Afrika" lockar stöd bland "ut ur Afrika"-anhängarna. De överväger i princip inte alternativa förklaringar.
Avslutningsvis övervägandet av figuren från artikeln (Scozzari et al, 2012) är det värt att notera det faktum att den allmänt accepterade ”icke-afrikanska” kombinerade haplogruppen ST (nedre raden i figuren) inte kommer ut ur haplogrupp B med dess underkläder, vare sig vi kallar det afrikanskt eller inte. Det kan ses att ST-haplogruppens evolutionära väg går förbi alla "afrikanska" haplogrupper, även om de alla innehåller europeiska eller andra förfäder. Om vi flyttar uppåt på tidsskalan (det vill säga från vänster till höger), lämnar mutationsvägen för CT-haplogruppen den huvudsakliga "stammen" av utvecklingen av den mänskliga Y-kromosomen, det vill säga under "svansen" som anges i figuren längst upp till vänster, passerar genom A0-T haplogruppen (ingen information om att den ska vara "afrikansk" - nej), sedan genom haplogrupp A1 (samma sak, det finns ingen information om att den ska vara "afrikansk"), sedan genom haplogrupp A1b, också icke-afrikansk, sedan genom haplogrupp VT, och blir haplogrupp ST. Alla tre "afrikanska" grenar (A0, A1a, A1b1) på denna väg går åt sidan genom motsvarande grenar och gafflar.
Vi uppehöll oss så detaljerat vid detta eftersom en sådan eller liknande analys aldrig har genomförts i den akademiska litteraturen, och särskilt i populärlitteraturen. Vanligtvis visas ett träd, som på bilden från artikeln (Scozzari et al, 2012), och smattret får höra att det "vittnar om mänsklighetens utträde från Afrika." Ingen förklaring ges. Svaret är redan klart. Ibland når det absurditet - det rapporteras att haplogrupperna BT och ST kommer från haplogrupperna A1b, eller A1, eller A0-T, vilket betyder att dessa är afrikanska haplogrupper, eftersom de har indexet "A". Det vill säga att ersättningen av begrepp har gått så långt att helt konventionella namn tas som bevis på tesen. Det faktum att dessa haplogrupper lika gärna kunde kallas X, Y, Z eller W kommer inte längre att tänka på. Eftersom "A" betyder Afrika råder det ingen tvekan.
Så det finns ingen motsägelse mellan figuren från artikeln (Scozzari et al, 2012) och diagrammet ovan; de visar samma mönster av evolutionär utveckling av haplogrupper, nämligen divergensen i "afrikanska" och "icke-afrikanska" grenar. Den enda skillnaden mellan dem är att figuren från artikeln (Scozzari et al, 2012) visar mer i detalj undergrupperna av haplogrupperna A (”afrikansk”) och B, och diagrammet visar de ”icke-afrikanska” haplogrupperna BT. En annan sak är att den första bilden är byggd med hänsyn till den kronologiska skalan, och den andra inte. För att visa likheterna, låt oss placera båda haplogruppträden vertikalt, i samma riktning.
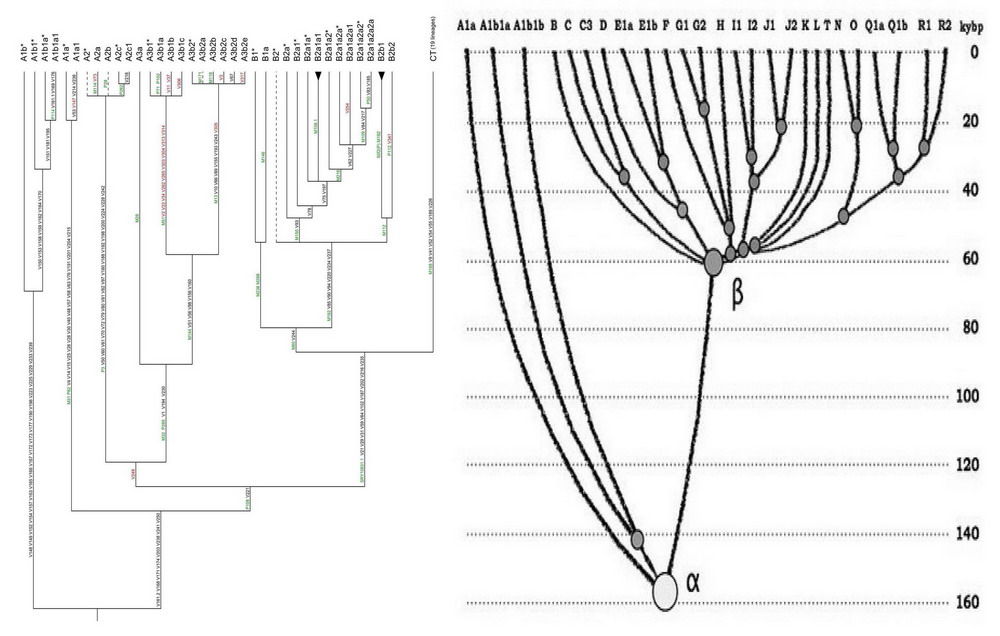
Jämförelse av haplogruppsträd och subklader av den "afrikanska" gruppen (den vänstra delen av båda träden) och den "icke-afrikanska" gruppen (en rad av den kombinerade ST haplogruppen på det vänstra trädet och en buske av BT haplogrupper på det högra trädet ). Det vänstra trädet (Scozzari et al) publicerades i november 2012, det högra trädet (Klyosov, Rozanskii) publicerades i maj 2012. Det vänstra trädet visar att serien av "afrikanska" haplogrupper avviker tre gånger i följd från de icke-afrikanska, och att den icke-afrikanska CT DNA-linjen (vertikal linje till höger) inte härstammar från de "afrikanska". Det högra trädet visar samma uppdelningar av "afrikanska" haplogrupper från icke-afrikanska (haplogruppbuske till höger), och indikerar att divergensen av trädet i afrikanska och icke-afrikanska haplogrupper inträffade för ungefär 160 tusen år sedan. Bilden är klickbar.
Båda träden visar inte den nyligen upptäckta haplogruppen A00, som är minst 200 tusen år gammal. Det visas i följande figur, tillsammans med ändringar i nomenklaturen (enligt ISOGG, 2013).
Så bilden som helhet har blivit tydligare. Det finns ingen motsägelse mellan trädet för mänsklighetens Y-kromosomala haplogrupper som erhållits från studiet av haplotyper av haplogrupper från A till T (Klyosov och Rozhanskii, 2012b, maj 2012) och trädet som erhållits från den genomiska studien av Y-kromosomen (Scozzari et al, 2012, november 2012) , nr. Alla dessa data, liksom andra, visar djup mutationell divergens mellan afrikanska och icke-afrikanska linjer (haplogrupper, subklader), och avslöjar inte det "afrikanska" ursprunget till den anatomiskt moderna mänskligheten. Istället visar uppgifterna skillnaden mellan afrikanska och icke-afrikanska DNA-linjer för omkring 160 tusen år sedan.
En logisk och förbryllande fråga uppstår: varför, med alla dessa data, fortsätter författarna till studierna att skriva att mänskligheten lämnade Afrika relativt nyligen, under de senaste 50-100 tusen åren? På vilken nivå av faktamaterial eller tolkning sker nedbrytningen? Att svara på denna fråga verkar inte mindre viktigt än att få svaret att mänskligheten inte lämnade Afrika.
Låt oss titta på den nämnda artikeln av Scozzari et al (2012). När dök uttrycket om människans afrikanska ursprung upp där? Vad är det baserat på?
Denna fras förekommer redan i det andra stycket i inledningen till artikeln och kommunicerar att CT-haplogruppen är resultatet av ett "nyligen utträde från Afrika." Till stöd för detta ges en länk till en artikel från 1000 Genomes Project-konsortiet med titeln "Map of Variation in the Human Genome" (Nature, 2010), där det inte finns några ord alls om utträdet från Afrika, inte heller om CT-haplogruppen. Förstår du vad problemet är? Förespråkare av konceptet "ut ur Afrika" måste ständigt fångas i handen, och detta har pågått i mer än 20 år. Några fler stycken nedan - igen om "utträdet från Afrika", redan haplogrupp C, och ingen referens alls.

Diagram över utvecklingen av den moderna mänsklighetens haplogrupper med tillägg av den nyligen upptäckta haplogruppen A00, och med ersättningen av den föråldrade nomenklaturen från 2012 med 2013 års nomenklatur. På den horisontella axeln finns huvudhaplogrupperna för mänsklighetens Y-kromosom, på den vertikala axeln är den absoluta tidsskalan. Den gemensamma förfadern till alfa-haplogruppen (haplogrupp A1b i den nuvarande klassificeringen) levde för cirka 160 tusen år sedan, den gemensamma förfadern till beta-haplogruppen (eller förfader för haplogrupperna B till T) levde för 64 ± 6 tusen år sedan (från artikeln av A.A. Klyosov och I.L. Rozhanskii, Advances in Anthropology, 2012b).
Låt oss följa artikeln (Scozzari et al, 2012) vidare. Haplogrupp A1b beskrivs (den översta linjen i figuren hämtad från den här artikeln, som var den första att avvika från resten av trädet, och kallas haplogrupp A0 under den nya nomenklaturen). Den flyttar sig också till vänster om trädet på det uppdaterade diagrammet över utvecklingen av haplogrupper av den moderna mänskligheten för ungefär 180 tusen år sedan. Det rapporteras att väldigt få personer med denna mutation (P114) har hittats, endast tre från Kamerun, en av dem i detta arbete. Min kommentar är mycket bra, jag har personligen inga stora tvivel om att haplogrupp A0 och dess grenar är afrikanska. Men vi härstammade inte från dem, som trädet visar.
Vidare rapporterar författarna att de i Niger hittade två personer i haplogrupp A1a - den andra raden i den övre delen av figuren från artikeln (Scozzari et al, 2012), också "afrikansk". Min kommentar - och det är inga problem med detta. Det finns inga icke-afrikanska ättlingar från dem heller, enligt samma figur.
Därefter är haplogrupp A2, det vill säga enligt den nuvarande nomenklaturen A1b1a, den tredje raden i figuren från artikeln. Författarna rapporterar att talare av denna haplogrupp nästan alla talar snappande språk i södra Afrika och är också centralafrikanska pygméer. Författarna hittade tre bärare av denna haplogrupp i Sydafrika. Min kommentar är jättebra, inga problem, det här är en helt afrikansk linje, och icke-afrikaner härstammar inte från den, som samma bild visar.
När det gäller haplogrupp A3, det vill säga enligt den nuvarande nomenklaturen A1b1b-M13, hittade författarna tio bärare av denna haplogrupp - i Etiopien, Kenya och Sydafrika. Ytterligare 28 personer, ägare till denna haplogrupp från europeiska länder och Saudiarabien, är listade i Haplogroup A-projektet, till vilket länken ges ovan. Men även om denna haplogrupp anses vara afrikansk, härstammar icke-afrikaner fortfarande inte från den, vilket framgår av figuren i artikeln (Scozzari et al, 2012). Författarna placerar haplogrupp B i Afrika söder om Sahara, såväl som i hela Afrika – centrala, östra och södra. Vi har redan visat ovan att haplotyperna för haplogrupp B är extremt avlägsna från de "afrikanska" och helt klart har ett annat ursprung, relaterat till icke-afrikanska haplogrupper. Men detta spelar ingen roll i det här fallet, eftersom de "icke-afrikanska" DNA-linjerna i haplogrupp ST inte kommer från haplogrupp B. De har en gemensam förfader med den - haplogrupp VT.
Och efter allt detta, vilken slutsats drar artikelförfattarna (Scozzari et al, 2012)? Du, läsaren, kommer att skratta, men slutsatsen är att ST-haplogrupperna kom från Afrika, och inte bara från Afrika i allmänhet, utan från dess nordvästra region. Det är där, enligt författarna, som "ursprunget till mänsklighetens Y-kromosommångfald" ligger. Hur var? Trots allt visar till och med en översiktlig blick på haplotypträdet att det inte finns några afrikanska källor på grund av ST. De är alla överst i diagrammet. Och så här. Det finns ett talesätt i Amerika om detta: "Förväxla mig inte med fakta, jag är redan klar."
Som läsaren redan har förstått, och förstått under lång tid, har "mänsklighetens utträde från Afrika" blivit en formell religion, baserad, som en religion borde vara, på tro, och argument där är nästan värdelösa.
Detta är också fallet i en annan relativt ny artikel (Cruciani et al, 2011), som redan har ”the origins of diversity in Africa” i titeln. På vilken vetenskaplig grund? Ja, allt är sig likt - de visade att afrikanska Y-kromosomala linjer är äldre än linjer från icke-afrikaner. Återigen enligt diagrammet ovan. Deras träd av haplogrupper är nästan detsamma som i figuren från artikeln (Scozzari et al, 2012), men med datum - för 142 tusen år sedan flyttade grenen A1b (A0 i den nya klassificeringen) bort från den evolutionära Y-kromosomala stammen, sedan för ungefär 108 tusen år sedan avgick gren A1a, sedan för 105 tusen år sedan, gren A2, sedan samma gren A3 för 105 tusen år sedan, som författarna anser vara afrikansk, eftersom de hittades i fyra afrikaner i vilka dessa haplogrupper fanns bestämt - och låt dem räkna, och först då, för 75 tusen år sedan, avgick VT-grenen och sedan, för 39 tusen år sedan, ST-grenen, redan allmänt erkänd som icke-afrikansk. Varken BT eller CT kommer från "afrikanska" linjer. Men eftersom författarna namngav förgreningspunkterna med bokstaven "A" (A1a-T, A2-T), som automatiskt tas som "afrikansk", så är detta "ursprung från Afrika". Denna typ av rättframt tänkande bland populationsgenetiker är slående.
Okej, det här är den manliga Y-kromosomen. Det är tydligt att motsvarande experimentella data inte visar någon utgång från Afrika. Det finns inga afrikanska SNP-mutationer vid basen av icke-afrikanska haplogrupper. Icke-afrikanska haplotyper är exceptionellt avlägsna från afrikanska. Som nästan alla källor medger är det ett enormt gap mellan dem, men författarna går inte längre. När de sluter ögonen upprepar de som ett mantra - "vi kom ut ur Afrika."
Korsvalidering av ett diagram med hjälp av snips (SNP)
Det finns ett annat tillvägagångssätt för att kontrollera topologin för trädet som presenteras i diagrammet precis ovan, med hjälp av klipp. Faktum är att under den evolutionära utvecklingen av Y-kromosomen ackumuleras praktiskt taget irreversibla mutationer i den, den så kallade SNP (Single Nucleotide Polymorphism), eller SNP. Ju längre övergången i diagrammet är, desto större är sannolikheten för en snip, desto fler av dem ackumuleras i Y-kromosomen (och i andra kromosomer, men i det här fallet överväger vi endast Y-kromosomen. Den längsta evolutionära linjen är haplogrupp A00, det är den mest arkaiska, därför bör bärare av denna haplogrupp avslöja det maximala antalet SNP:er. På andra plats när det gäller längd är haplogrupp A0, på tredje plats är linjerna i haplogrupp A (A1a). Avståndet mellan alfa- och beta-haplogrupper (det vill säga från A1b till BT) bör vara relativt små (jämfört med längden på linjerna A00 och A0), och sedan ackumuleras SNP:er redan under övergången från VT i linje med specifika, mer moderna haplogrupper .
Och så blev det, som en bekräftelse på diagrammet. Jag kommer nu att ge en lista med klipp för var och en av de listade haplogrupperna - å ena sidan skapa en sorts kompakt referensbok så att vem som helst kan arbeta med den, och å andra sidan för att inte bara begränsa sig i ord till ett visst antal som är svårt att verifiera. Det är sant att dessa siffror inte är slutgiltiga – nya klipp upptäcks då och då. Dessutom, eftersom SNP:er verkar oordnade, har vi att göra med statistik, och inte med absoluta, oföränderliga tal. Så listan nedan och antalet SNP:er för varje haplogrupp är under utveckling, även om de ger en allmän uppfattning.
Så - haplogrupp A00, den äldsta, den mest arkaiska, dess evolutionära linje i diagrammet är den längsta. Dessa snipper identifierades i Mbo-stammen (Mbo, med ryska bokstäver) i afrikanska Kamerun:
AF4, AF5, AF7, AF8, AF9, AF10, AF13, L990, L1086, L1087, L1088, L1091, L1092, L1094, L1096, L1097, L1100, L1102, L11043, L11043, L11043, L11043, L11043, L11043, L11043, L110 , L1110, L1111, L1113, L1114, L1115, L1117, L1119, L1122, L1126, L1131, L1133, L1134, L1138. 1, L1152, L1154, L1156, L1157 , L1158, L1159, L1160, L1161, L1163, L1233, L1234, L1236, L1284.
Totalt finns det 59 SNPs i haplogrupp A00. Eftersom åldern för haplogrupp A00 uppskattas till cirka 210 tusen år, kan det antas att en snipmutation inträffar i genomsnitt en gång vart 3600:e år.
Vidare, som vi redan vet, uppträdde haplogrupp A0-T på stammen av haplogruppsträdet (diagram ovan), haplogrupper A0 och A1 divergerade från den, den senare fortsatte med trädets stam. A0 bor nu huvudsakligen i Afrika. A0-T har inte en enda mutation från A00-listan. Det vill säga att A0-T inte härstammade från den afrikanska haplogruppen A00. Det finns ingen anledning att betrakta haplogrupp A0-T som afrikansk. Men hon är förfadern till den afrikanska linjen A0, och till oss, icke-afrikaner (vars förfäder passerade genom haplogruppen BT, sedan ST, och så vidare).
SNP:er för haplogrupp A0-T, det finns 32 av dem:
AF3, L1085, L1089, L1090, L1093, L1095, L1098, L1099, L1101, L1105, L1116, L1118, L1120, L1121, L1123, L11254, L12754, L12754, L12754, L12754, L127 1132, L1135, L1136, L1137, L1142, L1143, L1145, L1150, L1155, L1235, L1273
Haplogrupp A0 har följande klipp, det finns 51 av dem:
L529.2, L896, L982, L984, L990, L991, L993, L995, L996, L997, L998, L999, L1000, L1001, L1006, L1008. 018, L1055 . 22 5, V229, V233, V239
Som du kan se har haplogrupp A0 8 färre SNP än A00, det vill säga den är ungefär 30 tusen år yngre. Ja, åldern för haplogrupp A0 uppskattas till 180 tusen år, 30 tusen år mindre än åldern för haplogrupp A00.
I haplogrupp A1 har 21 SNP hittills identifierats:
L985, L986, L989, L1002, L1003, L1004, L1005, L1009, L1013, L1053, L1084, L1112, L1153.
Till skillnad från den broderliga haplogruppen A0 har bärare av A1 tydligen inte överlevt till våra dagar. Kanske fastnade de i den katastrof som ledde till flaskhalsen för världens icke-afrikanska befolkning. För att uppskatta den totala livslängden för haplogrupperna A0-T, A1 och A1b (den senare har bara två SNP-mutationer, P108 och V221), bör vi addera antalet SNP-mutationer, vi får 32+21+2 = 55 mutationer, vilket motsvarar ungefär 198 tusen år.
Slutligen, i BT-haplogruppen finns det 30 SNP-mutationer:
L413, L418, L438, L440, L604, L957, L962, L969, L970, L971, L977, L1060, L1061, L1062. V64, V102, V187, V202, V216, V235
Detta ger 108 tusen års evolution av beta-haplogruppen från tidpunkten för divergens från afrikansk haplogrupp A1b1 (160 ± 12 tusen år sedan) till passagen av befolkningens flaskhals för 64 ± 6 tusen år sedan. Detta är de saknade 108 tusen åren på diagrammet (avståndet mellan alfa- och beta-haplogrupperna).
Det viktiga är att dessa saknade årtusenden i utvecklingen av icke-afrikanska DNA-genealogiska linjer reproduceras både i analysen av haplotyper (på grundval av vilken diagrammet ovan konstruerades) och i analysen av snipmutationer. Detta är en korskontroll av diagrammet. Varför vi inte ser dessa försvunna människor i Eurasien är okänt. Å andra sidan är skelettrester kända Homo sapiens med en datering mellan 160 och 60 tusen år gammal, upptäckt i Mellanöstern, men deras haplogrupp har inte verifierats. Om haplogrupp BT visar sig ha någon av mutationerna som listas ovan kommer mysteriet äntligen att lösas.
Icke-afrikanskt mtDNA härstammar inte från afrikanskt
Vad säger data om kvinnligt mtDNA om detta? Tja, säger anhängare av "utträdet från Afrika", det är ännu tydligare där. Icke-afrikanska mtDNA är "alla härledda från afrikanska." Är det så?
Låt oss ta en titt på en ny artikel av den berömda genetikern D. Behar (Behar et al, 2012), där en grundläggande revidering av naturen hos mtDNA-sekvensrepresentation utfördes. Följande figur visar att i början av det evolutionära trädet av mtDNA för "den moderna människan" finns det en divergens av haplogrupper till L0 (gren till höger) och haplogrupper L1-L6 (gren till vänster), från vilka alla efterföljande haplogrupper uppstår därefter. Haplogrupp L0 representerar faktiskt en stor serie av femtio forntida afrikanska haplogrupper, främst i Afrika söder om Sahara - bland Khoisan-befolkningen i Sydafrika, men även i Etiopien och Tanzania (Östafrika), i Moçambique (Sydöstra Afrika) och bland pygméer . Allt annat mtDNA kommer, vilket är accepterat av de flesta experter, från haplogrupp L3, som uppskattas vara cirka 60-70 tusen år gammal, det vill säga samma som BT-haplogruppen i Y-kromosomen. Det vill säga, det är troligt att haplogrupp L3 inte lämnade Afrika, utan tvärtom kom till Afrika tillsammans med Y-kromosombärare, till exempel haplogrupp VT. Vilken slutsats drar Behar et al.? Naturligtvis kom "mänskligheten ut ur Afrika." Den djupa divergensen mellan den afrikanska haplogruppen L0 och de andra diskuteras inte ens i deras artikel, även om det från data i artikeln är omedelbart klart att den afrikanska haplogruppen L0 inte är förfäder till alla andra mtDNA-haplogrupper.

Schematisk representation av mänskligt mtDNA, som visar mutationsförhållandet mellan Neanderthal mtDNA (vänster) och Homo sapiens mtDNA (höger). I ovalen till vänster och höger står förkortningarna RNRS och RSRS för "rekonstruerad neandertalreferenssekvens" respektive "rekonstruerad mänsklig referenssekvens". Från arbetet (Behar et al., 2012). Notera den skarpa divergensen i början av den mänskliga referenssekvensen (höger), kedjan av mutationer för den afrikanska haplogruppen L0 (nederst till höger) och kedjan för alla andra mtDNA-haplogrupper. Bilden är klickbar.
Faktum är att skillnaden mellan den afrikanska haplogruppen L0 (åldern 150-170 tusen år) och de initialt icke-afrikanska haplogrupperna L1-L6, med den efterföljande ankomsten av haplogruppen L3 (åldern 60-70 tusen år) i Afrika visar sig vara praktiskt taget sammanfaller med divergensen av de Y-kromosomala haplogrupperna i den afrikanska serien A (ålder 160 tusen år) och initialt icke-afrikansk VT (ålder 64 tusen år) med den efterföljande ankomsten av haplogrupp B till Afrika.
Så när det gäller mtDNA visar sig konceptet att "gå ur Afrika" vara helt onödigt, och är faktiskt byggt på sand. Beskrivningar av mtDNA i akademiska artiklar och referensböcker är fulla av orden "möjligt", "sannolikt" och "föreslagit", vilket i huvudsak betyder att det inte finns några data och att allt är baserat på spekulationer. Ett problem är att alla dessa antaganden alltid tolkas i bara en riktning - "utträde från Afrika."
Samtidigt ackumuleras fler och fler bevis för att i det gamla förflutna förekom migrationer av befolkningar till Afrika många gånger. En artikel har just publicerats i tidskriften Nature (Hayden, 2013) som rapporterar två migrationer från Eurasien till Khoisan-stammarna i Sydafrika, den ena för 3 tusen år sedan till östra Afrika, den andra, dess fortsättning - för 900-1800 år sedan till Sydafrika . Det rapporteras inte vilka haplogrupper migranterna tog med sig. Men det råder ingen tvekan om att de dramatiskt ökade den "genetiska mångfalden" hos Khoisan, som anses vara den högsta i Afrika. Det andra viktiga inslaget i detta meddelande är att det indikerar möjligheten till migrationer "till Afrika", vilket det dock inte kunde råda några tvivel om. Det är inte klart varför anhängare av "mänsklighetens utträde från Afrika" håller fast vid sitt ensidiga alternativ med sådan envishet. Uthålligheten avtar dock, och nu välkomnar Sarah Tishkoff från University of Pennsylvania, en av de mest aktiva förespråkarna för "out of Africa", redan nya data "till Afrika" och kallar det "förnuftigt", eftersom " både arkeologisk och språklig forskning” (Naturen, 29 augusti 2013, s. 514).
Herpesvirus migrationer rapporteras felaktigt som
påstås bekräfta "mänsklighetens utträde från Afrika"
Berättelsen om den senaste sensationella publikationen "Using phylogenetics of the herpes simplex virus genome to trace forntida mänskliga migrationer" är en indikation på hur det initiala antagandet om den påstådda framväxten av modern mänsklighet från Afrika förvränger slutsatserna i en vetenskaplig publikation. Låt mig påminna dig om att artikeln jämförde 31 genomiska sekvenser av herpes simplex-virus i populationer från Östafrika (Kenya), Östasien (Kina, Sydkorea, Japan), Nordamerika (USA) och Europa (Storbritannien), och fann att den konstruerade fylogenetiken divergerar trädet i sex kluster. Dessa kluster motsvarade följande populationer:
I – av 10 prover var 7 från Seattle, ett "från USA", ett från San Francisco och det nedersta (nummer 17) från Skottland.
II – av 6 prover var ett från Kina, ett från USA (Houston), två från Sydkorea och två från Japan.
III – två prover från Kenya.
IV – tre prover från Kenya.
V – sju prover från Kenya.
VI – två prover från Kenya.
Författarna till artikeln "rundade" uppgifterna och rapporterade att det första klustret "förenar Nordamerika och Europa", det andra - Östasien, resten - "Östafrika". Ursprunget till de amerikanska virusproverna anges inte i artikeln, det vill säga det är oklart om de erhållits från amerikanska indianer eller till exempel ättlingar till anglosaxarna, men eftersom författarna föreslog att provet från Houston, som föll ur även den rundade versionen, som hamnade bland de östasiatiska virusen, kunde tillhöra American Indian, blir det tydligt att de återstående nordamerikanska proverna med största sannolikhet kommer från ättlingar till européer. Författarna drog dock slutsatsen att dessa kluster återspeglar "globala migrationer av (forntida) människor" och stödjer framväxten av den moderna mänskligheten från Afrika (stödjer ... "ut ur Afrika" teorin om mänsklig evolution). Det främsta skälet är, som vanligt, argumentet för "mångfald", nämligen att "virus av östafrikanskt ursprung har den största mångfalden och bildar fyra av de sex klustren."
Låt oss, som vi gjorde ovan, titta på var denna "mångfald" kommer ifrån. Och det framgår igen av skillnaderna mellan befolkningar i olika riktningar - vissa går till Afrika, andra lämnar inte. Men återigen härstammar de inte från varandra, varje gång härstammar de från en mer gammal gemensam förfader, som kanske inte har bott i Afrika. Det vill säga att de situationer som beskrivits tidigare i den här artikeln upprepas igen. Data som visas indikerar inte något "afrikanskt ursprung".

Fylogenetiskt träd som visar dynamiken hos stammar av herpes simplex-viruset HSV-1 och dess kaskaddivergens i sex grenar, räknat i (Kolb et al., 2013), som enligt samma författare följer en viss geografi: gren I - Europa / Nordamerika (stam 17 - från Glasgow, resten - Seattle, San Francisco, "USA"), gren 2 - Östasien (Kina, Sydkorea, Japan och Houston), grenar III, IV, V och VI - Östafrikanska (alla från Kenya).
Låt oss titta på diagrammet nedan. Till vänster är den första avvikelsen från en gammal gemensam förfader. Bortsett från de virus som nu finns i kenyaner (de två översta proverna, kluster VI). Inga icke-afrikanska virus kommer från dem.
Nästa - en annan gaffel, en annan divergens av populationer. Det kenyanska viruset flyttar återigen åt sidan, med index E07 i diagrammet ovan. Återigen, icke-afrikanska virus kommer inte från det. Dess kombination med resten av de kenyanska proverna i kluster IV är ett misstag av författarna; det finns inget gemensamt kluster där.
Nästa är ytterligare två befolkningsavvikande gafflar, och varje gång flyttar den kenyanska gruppen (kluster IV och III), som européer och asiater inte kommer från, bort. Vid den sista divergensen är det uteslutande den icke-afrikanska gruppen som flyttar åt sidan. Återigen, det kommer inte från afrikaner.
Den senaste skillnaden är mellan europeiska och östasiatiska (med vissa undantag) härstamningar av herpesvirus, kluster I och II. Amerika har tydligen ingenting med det att göra, det är inte de gamla invånarna i Amerika, utan relativt nya immigranter från Europa.
I en kommentar till tidningen sa huvudförfattaren Curtis Brandt, professor i medicinsk mikrobiologi och oftalmologi, att resultaten var "häpnadsväckande" och fortsatte med att säga att "vi fann att alla afrikanska isolat bildade ett kluster, alla virus från Fjärran Östern , Korea, Japan, Kina bildar tillsammans ytterligare ett kluster, och alla virus från Europa och Amerika, med ett undantag, bildar ytterligare ett kluster." I allmänhet, trots de uppenbara sträckorna, är detta sant. Men visar detta "den moderna människans afrikanska ursprung"? Uppenbarligen inte, som förklarats ovan.
I en annan kommentar delade Dr. Brandt helt öppet: "Vi hittade exakt vad molekylärgenetiker som studerar det mänskliga genomet berättade för oss, nämligen var människor kom ifrån (Afrika - AK) och hur de divergerade över planeten." Detta är huvudproblemet i denna och liknande studier. De fick veta (felaktigt), de tog det som en vägledning till handling och hittade exakt detta.
Artikeln ger ytterligare en illustration av hur data skräddarsys för det önskade "utträdet ur Afrika." Så här beräknade författarna tiderna för denna "exit", baserat på de som antogs för 50 tusen år sedan, och som faktiskt aldrig visades. Detta är "konsensusdata". Faktum är att olika författare ger datum från 27 till 200 tusen år sedan, som regel, igen utan beräkningar, men av någon anledning ser siffrorna för 50 eller 70 tusen år sedan attraktiva ut för dem. Nyligen började emellertid "konsensus" att skifta till 100-140 tusen år sedan, men 50 eller 70 tusen år fortsätter att ges av tröghet. Så författarna till artikeln som diskuteras använde dessa spekulativa datum som basdatum. Låt oss se vad författarna fick ut av detta. Detta är ganska lärorikt.
Som författarna till artikeln rapporterade finns det i litteraturen tre signifikant olika värden för mutationshastighetskonstanter för herpes simplex-virus och andra herpesvirus, lika med 3x10 -9, 18,2x10 -9 och 30x10 -9 mutationer per nukleotid och år. Att använda dem för att beräkna tiden för virusets divergens bland de mänskliga populationerna som beskrivs ovan skulle ge en 30-faldig tidsskillnad. Det är uppenbart att författarna inte var nöjda med sådan osäkerhet, och de beslutade att faktiskt justera tidpunkterna för den initiala divergensen av populationerna till "konsensus"-tiden för "utträde från Afrika", och i det första skedet av beräkningar att justera till den beräknade tidpunkten för divergensen mellan de europeiska och asiatiska befolkningarna, som de tog för 23-45 tusen år sedan, med hänvisning till fyra litterära källor om detta ämne. Efter att ha tagit ett medelvärde för dessa värden tog författarna 34 000 ± 10 500 år sedan som ett "referensvärde". Det är sant att i stället för "europeiskt" nämnde författarna ständigt "europeiskt/nordamerikanskt", även om det är tydligt att det i huvudsak är europeiskt - ett prov av viruset är från Skottland, alla andra (mest från Seattle) är troligen ättlingar av invandrare från Europa. I vilket fall som helst är dessa siffror underskattade, eftersom separationen av haplotyperna i Europa och Asien skedde senast för 55-60 tusen år sedan, de äldsta benresterna av "anatomiskt moderna människor" i Europa går tillbaka till 45 tusen år sedan, aboriginerna kom till Australien senast för 50 tusen år sedan år sedan, men allt i artikeln är så ungefärligt - både dateringen och mutationshastigheten för viruset - att detta inte kan tas på allvar. Eftersom författarnas tider är mycket underskattade borde mutationshastigheterna ha varit mycket högre. Och så blev det - vijusterad på detta sätt visade sig vara mycket snabbare än de kända (mer exakt, litterära) värdena, nämligen 134x10 -9 mutationer per nukleotid och år, med övre och nedre gränser på 214x10 -9 och 74,8x10-9, respektive.
Med hjälp av denna justerade mutationshastighetskonstant uppskattade författarna att den ursprungliga divergensen av viruset inträffade för 50,3 ± 16,7 tusen år sedan och fastställde att detta motsvarar "mänsklighetens utträde från Afrika." En något korrigerad datering av divergensen mellan européer och asiater, enligt författarnas beräkningar, visade sig vara 32,8 ± 10,9 tusen år sedan, och tidpunkten för divergensen för det enda kinesiska provet och det enda provet från Texas visade sig vara 15,76 ± 5,3 tusen år sedan, som författarna tillskrev bosättningen i Amerika, "som ägde rum vid den tiden." period". Kommentarer här är helt enkelt onödiga.
Allt detta låg till grund för författarnas deklaration att " det visas för första gången att fylogenetiska data om herpesvirus bekräftar mänsklighetens utträde från Afrika" Som faktiskt visas ovan har detta ingenting att göra med att "mänskligheten lämnar Afrika." Dessutom överstiger uppskattningen av mutationshastighetskonstanten för herpesviruset, utförd av författarna på basis av mer än grova uppskattningar, tre andra litteraturvärden med 4,5 - 45 gånger. Författarna utförde inga korsvalideringstester för att verifiera den erhållna mutationshastighetskonstanten. Det faktum att författarna erhöll, baserat på sin konstant, att herpesvirusen HSV-1 och HSV-2 divergerade för 2,184 ± 0,753 miljoner år sedan (observera den givna "precisionen" på tre decimaler!) betyder ingenting, det skulle kunna har hänt där för 20 miljoner år sedan med samma framgång, och det skulle inte ha sagt något heller - det kunde ha hänt i makaker till exempel. Om, trots allt, de litterära uppgifterna är mer korrekta, så kunde den initiala divergensen av herpesviruset ha inträffat inte för 50,3 tusen år sedan, utan för 220 tusen år - 2,2 miljoner år sedan, och kunde också tolkas väl inom ramen för människan Evolution. Så författarnas ursprungliga experimentella data är av otvivelaktigt värde, men manipulationer, slutsatser och tolkningar saknar värde. Tyvärr är det så vetenskap ofta "görs" idag, särskilt inom området för populationsgenetik.
Antropologiska data och datering
Under de senaste åren har en intressant situation uppstått när många antropologer uttrycker tvivel om "människans afrikanska ursprung", men pekar på genetiker som säger att vi inte har något svar, antropologiska data är motsägelsefulla, men genetiker hävdar att de vet för säkert från Afrika, så hur kan vi argumentera? Och de genetiker som förstår att genetisk data är byggd på sand, eller mer exakt, på alltför fria (fantasi-)tolkningar, nickar åt antropologer att, de säger, vi förstår att genetisk data är svag och ofta helt enkelt felaktig, men antropologer hävdar att från Afrika, och deras dating pekar på detta, så hur kan vi argumentera? Det betyder att allt stämmer hos oss.
Låt oss titta på påståendena att den anatomiskt moderna människan (ACH) påstås definitivt ha sitt ursprung i Afrika, och det var ungefär 40-50 tusen år sedan, eller 100, eller 150 eller 200 tusen år sedan. Låt oss komma ihåg att en ASP är någon som inte har märkbara arkaiska antropologiska egenskaper. Först kommer vi att skissa en kort beskrivning av situationen, sedan kommer vi att demonstrera detta med hjälp av specifikt material. För att uttrycka det riktigt kort har vi fem huvudpunkter:
(1) alla afrikanska fynd av forntida ben upp till omkring 36 tusen år sedan visar märkbara arkaiska drag;
(2) ofta är forntida ben så fragmentariska att det helt enkelt är omöjligt att återskapa ens en minimal antropologisk bild från dem;
(3) ofta är den antropologiska bilden av ben i Afrika och utanför Afrika väldigt lik, och därför kan man inte säga om det var en utgång från Afrika eller en ingång till Afrika;
(4) ofta finns det inga benrester alls, och uttalanden om "anatomiskt moderna människor" görs på grundval av platser och stenredskap, även om de mycket väl kunde ha lämnats av arkantroper, det vill säga arkaiska människor som inte tillhör arten "Homo sapiens";
(5) Dateringen av forntida ben är ofta så tveksam att få människor tar dem bokstavligt eller till och med på allvar.
Låt oss börja med den sista. Tyvärr fungerar radiokoldatering ibland större än för cirka 40 tusen år sedan nästan inte, och det senaste dateringsrekordet är 60 tusen år sedan. Anledningen är enkel - halveringstiden för den radioaktiva isotopen 14 C är 5730 år, det vill säga 40 tusen år är sju halveringstider och 60 tusen år är mer än tio halveringstider. Metoden bygger på att mäta förhållandet i biologiska prover av halten av den stabila isotopen 12 C (och lite 13 C, nästan hundra gånger mindre jämfört med halten 12 C) och radioaktivt 14 C (med dess initiala halt i mängden en tiomiljarddels procent), som med tiden sjunker, med samma halveringstid. Under 60 tusen år minskar dess innehåll från de ursprungliga 10 -10% med 2 10 gånger, det vill säga ytterligare 1024 gånger. Moderna enheter upptäcker inte längre sådana strålningsnivåer, åtminstone inte de enheter som används av arkeologer. Detta är ungefär 1 klick per timme per gram kol som testats. Den normala bakgrunden är mycket högre.
Detta öppnar för "vida möjligheter" för misstag, och inte bara misstag. Låt oss minnas den sensationella (i snäva kretsar) berättelsen om degradering och skandalöst avskedande av den tyske antropologen Reiner Protsch för att systematiskt, som det visade sig, förfalska dateringen av gamla (och inte alls antika) ben. Efter kontroll visade sig Protschs datum för 36 000 år sedan vara 7 500 år sedan, hans datum för 21 300 år sedan visade sig vara 2 300 år sedan och ett skelett han daterade till för 29 400 år sedan visade sig vara kvarlevorna av en man som dog 1750, 255 år före mätningarna (tidningen The Guardian) . Den berömda internationella tidskriften "Archaeology" skrev också om detta.
Det finns många liknande historier inom arkeologin, men även om vi kastar bort uppenbara förfalskningar, som det förstås bara finns ett fåtal av, så är sannolikheten för fel i alla fall stor. Särskilt när man verkligen vill göra saker uråldriga och därmed gå till arkeologins historia, och till och med mänsklighetens kultur som helhet. Det måste sägas att särskilt forntida biologiska fynd dateras med andra metoder, såsom argon, vad gäller 40 Ar/ 39 Ar-innehållet.
Sammantaget finns det inga antropologiska eller arkeologiska bevis för "den moderna människans ursprung i Afrika", och det finns inte heller några bevis för att de sten "verktyg" och "industrier" som finns i Europa eller Eurasien i allmänhet kom från områden söder om Sugars. Alla påståenden om upptäckten av skelettrester av "anatomiskt moderna människor" för mer än 50 tusen år sedan, och ännu mer för mer än 150 tusen år sedan, och särskilt söder om Sahara, är helt enkelt förvrängda eller felaktiga från början. En ganska omfattande recension av detta ämne kommer snart att publiceras av den välrenommerade australiensiske antropologen Robert Bednarik (kommande, Advances in Anthropology).
Frånvaron av sådana fynd söder om Sahara bevisas av ett antal verk, till exempel (Grine et al, 2007; Grine et al, 2010). Alla kända fynd av ben från en sådan forntid har uppenbara arkaiska drag, som börjar med Omo Kibish 1 (för 195 tusen år sedan, Etiopien, fragment av kranialben hittades, få ansiktsben), Omo-2 (visar ett antal primitiva, arkaiska drag ), Herto (154-160 tusen år sedan, en mycket arkaisk benstruktur, mycket annorlunda än ASC), i allmänhet är alla skelettrester från 100-200 tusen år sedan, och faktiskt före 35 tusen år sedan, arkaiska ( Rightmire, 2009). Många har inga bevarade ansiktsben alls. Till och med Hofmeyr-skallen från Sydafrika, från 36 tusen år sedan, har arkaiska drag (Grine et al, 2007; Rightmire, 2009; Tattersall, 2009).
Den amerikanska antropologen Rightmire rapporterade: " varken Herto-fossilerna eller andra från sent Pleistocen, som Klasiesfloden i Sydafrika eller Skhul/Qafzeh i Israel, har paralleller med moderna befolkningar. Deras skallar är robusta, och först från ~35 000 år sedan börjar människor med modern anatomisk morfologi dyka upp"(Rightmire, 2009). Han tror att "anatomiskt moderna människor" utvecklades i Afrika, även om processen är "dåligt förstådd". Han upprepas av Michael Hammer (Hammer et al, 2011) – " Fossila homininer som visar en kombination av arkaiska och mer moderna egenskaper finns konsekvent i Afrika söder om Sahara och Mellanöstern fram till för cirka 35 tusen år sedan" Således är de ständiga referenserna från anhängare av konceptet "ut ur Afrika" att benrester från anatomiskt moderna människor med datum mellan 160 och 200 tusen år sedan upptäcktes i Afrika falska. Manipulation och distorsion fortsätter även här.
Michael Hammer, på senare tid en aktiv förespråkare för konceptet "ut ur Afrika", började uttrycka tvivel om "ut ur Afrika", men började uttrycka detta redan 2013, när det stod klart att situationen med "utanför Afrika". Afrika” var extremt förvirrad. Hammer avslutar sin artikel i Scientific American (maj 2013) med följande ord: " Många knutar förblev upplösta. Men en sak är klar - den moderna människans rötter går inte bara till en enda förfädersbefolkning i Afrika, utan till befolkningar i den gamla världen"(det vill säga Europa eller Eurasien - AK).
Och detta verkar ganska rimligt. När vi studerar den antika världens historia möter vi ständigt mobila migrationer, inklusive mycket långdistansmigrationer. Om människan, enligt anhängarna av "out of Africa", nådde Australien på bara 10 tusen år, så är det svårt att föreställa sig att han satt stilla i 200 tusen år och inte spred sig över hela världen, inklusive till och från Afrika, och många gånger om. Hur det gick till att konceptet med ett ensidigt "utträde från Afrika" så aggressivt påtvingades samhället och tog tag i det så snabbt borde väcka varningsklockor – både när det gäller det aggressiva, tvingande påtvingandet och när det gäller sådana viljesvaga allmänhetens uppfattning. Och det som borde vara särskilt alarmerande är att det inte fanns och finns inga tillförlitliga skäl för att "gå ur Afrika."
För att sammanfatta, upprepar vi ännu en gång - våra förfäder lämnade inte Afrika under de senaste 200 tusen åren. Med andra ord, de som kom ut från Afrika födde inte den moderna icke-afrikanska mänskligheten. Detta visar hela komplexet av data som erhållits - genetik, antropologi, arkeologi och DNA-släktforskning. Faktum är att inte bara under de senaste 200 tusen åren, utan också tidigare. En studie av DNA från fossila Nenderthal-ben visade närvaron av melanokortinreceptorn (MCR1), och i en variant som specificerar ljus hud och rött hår (Lalueza-Fox et al, 2007). Författarna tror att neandertalarna hade ungefär samma hårfärg som moderna européer, allt från mörkt till blont. Dessutom har inga indikatorer hittats på att neandertalare var negroider. Faktum är att inga spår av neandertalare har hittats i Afrika. Och eftersom våra närmaste förfäder var gemensamma för neandertalarna, eftersom neandertalaren är vår brorson, så hade "far" till neandertalaren och "bror" till vår "far" också med största sannolikhet ljus hud och bodde inte i Afrika. Det var någonstans i intervallet 300-600 tusen år sedan. Det förblir dock olöst hur våra ljushyade bröder, som vi divergerade med för ungefär 160 tusen år sedan, överlevde när de kom till Afrika, och hur de fick mörk hudfärg, men svaret kan också hittas inom genetikområdet , vid reglering av melaninbiosyntes. Men det är en annan historia.
Anatoly A. Klyosov,
professor, doktor i kemivetenskap
Gillade du artikeln? Dela länken med dina vänner!
115 kommentarer: Våra förfäder lämnade inte Afrika
Anatoly A. Klyosov säger:
Andrey säger:
Anatoly A. Klyosov säger:
Arsens säger:
I. Rozhansky säger:
Avstod Charles Darwin från sin teori om mänsklig evolution i slutet av sitt liv? Hittade forntida människor dinosaurier? Är det sant att Ryssland är mänsklighetens vagga, och vem är yetien - kanske en av våra förfäder, förlorad genom århundradena? Även om paleoantropologin - vetenskapen om mänsklig evolution - blomstrar, är människans ursprung fortfarande omgiven av många myter. Dessa är anti-evolutionistiska teorier, och legender genererade av masskultur, och pseudovetenskapliga idéer som finns bland utbildade och pålästa människor. Vill du veta hur allt "egentligen" var? Alexander Sokolov, chefredaktör för portalen ANTHROPOGENES.RU, samlade en hel samling liknande myter och kollade hur giltiga de är.
Någon hävdade att centrum för mänskligt ursprung är där de så kallade "primitiva raserna" nu lever. Det fanns anhängare av polygenism som trodde att var och en av de tre stora raserna - "vit", "svart" och "gul" - hade sina egna apaförfäder. Polygenister har alltså inte ett enda centrum: raser uppstod oberoende i olika delar av världen.
Efter upptäckterna av Pithecanthropus i Java, och sedan Sinanthropus i Kina, såg många, inklusive sovjetiska antropologer, det troliga centrumet för antropogenesen i Asien. ”Människans hemland var utan tvekan den eurasiska kontinenten. Här, i Centralasien, med all sannolikhet, där Gobiöknen nu ligger, uppstod den situation som ledde till att det första apfolket dök upp”, skrev den sovjetiske paleontologen A.P. Bystrov i mitten av 1900-talet.
De första upptäckterna av Australopithecus i Afrika möttes av fientlighet av många forskare - beror det på att den europeiska förfadern är mycket ädlare än någon afrikansk apa? Men låt oss inte förenkla det. I sin bok "Fossil Chronicles" försöker den amerikanske paleoneurologen Dean Falk lista ut: varför tog det 25 år för den vetenskapliga världen att känna igen "Child from Taung" - den första upptäckten av en australopithecus som Raymond Dart gjorde i Sydafrika? Falk citerar en annan berömd specialists ord, Philip Tobaios:
Upptäckten som beskrevs av Dart var före sin tid, eftersom dess innebörd inte kunde kopplas samman med enkla logiska steg med den tidens idéer. Taungs fördröjning av erkännande är inte unik, noterade Tobaios, eftersom samma historia hände med ett antal andra viktiga "för tidiga" upptäckter, såsom genetikens lagar eller penicillin. Tobayos listade ett antal principer för mänsklig evolution, allmänt accepterade 1925, som Taung utmanade. Bland dem var påståendet (nu erkänt som felaktigt) att Asien är mänsklighetens vagga; att ökningen i hjärnstorlek "föregick" hominidutveckling, vilket antyddes av Piltdown-fyndet; att de flesta av Taungs drag kunde förklaras av hans unga ålder, och att geo; Taungs logiska datering var för sent för en mänsklig förfader. ( Per. bil)
Det var först efter upptäckterna som Lewis Leakey och andra vetenskapsmän gjorde i Östafrika under andra hälften av 1900-talet som bilden började bli tydligare. Homo habilis upptäcktes i Olduvai (Tanzania), afrikanska erecti hittades, de äldsta sapiens från Omo (Etiopien) beskrevs, och slutligen spreds nyheten om upptäckten av Australopithecus Lucy i Hadar (Etiopien) över världen. Faktum för faktum, sten för tegel, banade den afrikanska versionen vägen till vetenskapligt erkännande.
Hittills har resterna av de äldsta hominiderna hittats i Afrika (med början från Sahelanthropus för cirka 7 miljoner år sedan). Den evolutionära kedjan av afrikanska fynd är den mest kompletta:
(Listan inkluderar inte hominider som förmodligen inte är direkt relaterade till våra anor.)
Ingen annan kontinent har en så detaljerad kedja; Det finns inte heller några extraafrikanska fynd av hominider äldre än 2 miljoner år – forntida människa dyker upp utanför den afrikanska kontinenten efter detta datum.
Det är viktigt att det var i Afrika som de äldsta människorna av den moderna typen och deras omedelbara förfäder upptäcktes. Så det kan hävdas att i Afrika uppstod som ett släkte Homo, och, mycket senare, vår art - Homo sapiens (se även myt nr 24).
Arkeologi säger samma sak. De äldsta stenverktygen som finns i Afrika (detta är platsen för Gona, Etiopien, för 2 600 000 år sedan). Alla arkeologiska fynd utanför Afrika är yngre än 2 miljoner år.
När det blev möjligt att bygga släktträd med hjälp av genetiska data, samlade genetiker DNA-prover från människor av olika raser och byggde ett släktträd av mänskligheten. Det visade sig att tidigare än andra skildes en gren som bara innehöll afrikanska grupper från stammen på detta träd. Dessutom visade det sig att den genetiska mångfalden hos människor minskar när de flyttar bort från Afrika. Detta är logiskt: en grupp gamla Homo sapiens, som en gång lämnade den afrikanska kontinenten, tog bara en del av den afrikanska genpoolen till Eurasien. Därefter upprepades denna situation många gånger - trots allt bosatte de sig som regel i små grupper, varje gång tog de bara med sig en bit av genpoolen.
För inte så länge sedan, 2011, kom ytterligare bevis på mänskligt afrikanskt ursprung från lingvistik. Nyzeeländsk psykolog och antropolog Quentin Atkinson analyserade ljudmångfalden hos 504 språk i världen och visade att ju längre från Afrika, desto lägre antal fonem - de minsta språkliga enheterna - i språk. Det antas att samma regel gäller för språk som för gener: fonetisk mångfald (liksom genetisk) är maximal i "ursprungscentrum". (Men riktigheten av forskningen bekräftas av vissa forskare, inklusive Svetlana Burlak, en specialist på språkens ursprung.
En genetisk studie av kaukasier fann att vita människor härstammar från en grupp på cirka 20 personer, med dubbelt så många män som kvinnor.
I oktober 1997 hölls en konferens om mänsklig evolution i Cold Spring Harbor (USA). De ständiga hänvisningarna till teorin om afrikanskt mänskligt ursprung och forskning om afrikanska och icke-afrikanska befolkningar föranledde en oroande kommentar från en deltagare om etiken i en sådan uppdelning av mänskligheten, förknippad med den allmänna opinionens känslighet i USA när det gäller rasfrågor. .
Den schweiziska genetikern Dr Andre Langani svarade kvickt på detta: istället för den planerade rapporten om genogeografi höll han en föreläsning om att vi alla är afrikaner, att döma av genetiska data, det finns helt enkelt asiatiska afrikaner, europeiska afrikaner och afrikanska afrikaner.
Idag följer de flesta forskare teorier om afrikanskt mänskligt ursprung och de tror att den framtida vinnaren i den evolutionära rasen uppstod i sydöstra Afrika för cirka 200 tusen år sedan och bosatte sig därifrån över hela planeten (Fig. 1).
Sedan människan kom ut ur Afrika tycks det vara en självklarhet att våra avlägsna afrikanska förfäder liknade de moderna invånarna på denna kontinent. Vissa forskare tror dock att de första människorna som dök upp i Afrika var närmare mongoloiderna.
Mongoloid ras har ett antal arkaiska drag, särskilt i tändernas struktur, som är karakteristiska för neandertalare och Homo erectus (Homo erectus). Populationer av mongoloidtyp är mycket anpassningsbara till olika levnadsförhållanden, från den arktiska tundran till ekvatoriska regnskogar, medan hos barn av den negroida rasen på höga breddgrader, med brist på vitamin D, utvecklas bensjukdomar, rakitis, dvs. snabbt. de är specialiserade på förhållanden med hög solinstrålning. Om de första människorna var som moderna afrikaner är det tveksamt att de skulle ha lyckats migrera runt jorden. Denna uppfattning ifrågasätts dock av de flesta antropologer.
Begreppet afrikansk härkomst är emot multi-regionalt ursprung koncept, vilket tyder på att vår förfäders art Homo erectus utvecklades till Homo sapiens oberoende på olika ställen runt om i världen.
Homo erectus uppträdde i Afrika för cirka 1,8 miljoner år sedan. Han tillverkade stenredskapen som paleontologer hittat och möjligen mer avancerade bambuverktyg. Men efter miljontals år finns det inga spår av bambu. Under loppet av flera hundra tusen år spred sig Homo erectus först över Mellanöstern, sedan in i Europa och till Stilla havet.
Bildandet av Homo sapiens på basis av Pithecanthropus ledde till samexistensen av senare former av neandertalare och framväxande små grupper av moderna människor under flera tusen år. Processen att ersätta en gammal art med en ny var ganska lång och därför komplicerad.
Intressant i dess ovanlighet Porshnev koncept, ett alternativ till darwinismen. Porshnev anser att de gamlas förändring i sättet att äta är den avgörande faktorn i evolutionen. Deras gradvisa övergång från att äta "från någon annans bord" med skrot och kadaver till att övervinna det interna förbudet "du ska inte döda!" och kannibalism. Porshnev identifierar tre grenar : rovdjur, manipulatorer och offer. Han extrapolerar deras utveckling till representanter för det moderna, heterogena mänskliga samhället. Han anser att rovdjurens ättlingar är den mäktiga – den rikaste klassen, ättlingar till manipulatorer – politiker, den byråkratiska klassen och offren – naturligtvis, fattiga människor. Svagheten med hans teori, enligt min mening, är att han anser att varje persons öde är förutbestämt, hans undervisning förnekar valfrihet, hans teori kan inte förklara människors övergångar mellan dessa tre klasser som äger rum i det verkliga livet.
IVMänniskans biologiska utveckling idag
Social information överförs genom ord under inlärning och bestämmer individens andliga utseende. Det skapas med den dominerande rollen av socioekonomiska faktorer - social bildning, nivå på produktionskrafter, produktionsrelationer, nationella särdrag och rp.
Människan som social varelse utvecklas snabbare än som biologisk varelse, därför finns det, trots civilisationens enorma landvinningar, inga betydande biologiska skillnader mellan en person som levde för tusentals år sedan och en person som lever nu.
En persons förmågor, talang, emotionalitet, dygder och laster beror på ärftliga anlag och den sociala miljöns handlingar. En persons genotyp ger möjlighet att uppfatta ett socialt program, och det fullständiga genomförandet av dess biologiska organisation är endast möjligt under villkoren i en social miljö.
Även om mutationsprocessen fortsätter, kommer den mänskliga biologiska evolutionen att fortsätta att sakta ner på grund av att det naturliga urvalet försvagas och dess artbildande funktion upphör. Men inom en art är fluktuationer möjliga: i kroppslängd (medeltida riddares rustning är liten för de flesta moderna européer), förändringar i ontogeneshastigheten (acceleration av tonåringar), etc.
Vitaliteten i det mänskliga samhället som helhet ökar, eftersom när civilisationen utvecklas och nationella och rasmässiga barriärer elimineras, säkerställs utbytet av gener mellan tidigare isolerade populationer, heterozygositeten ökar och möjligheten för manifestation av recessiva gener minskar.
I humaniseringsprocessen sker en minskning av fertiliteten, en förlängning av barndomsperioden, en avmattning i puberteten och en ökning av livslängden för en generation.
Medlen som styr mänsklig evolution är skydd mot effekterna av mutagena faktorer, utveckling av metoder för behandling av ärftliga sjukdomar, utveckling av mänskliga förmågor i barndomen och tonåren och skapandet av optimala förutsättningar för träning och utbildning, för att förbättra den kulturella hela samhällets nivå.
Och så såg vi människans naturliga ursprung. Vi såg att tidigare, för många tusen år sedan, var människan en del av naturen lika med resten. Men i evolutionsprocessen, steg för steg, separerade människan sig från den. Efter att ha börjat från att lämna skogen och hålla en pinne i sina händer har en man redan nått atombomber, men har fortfarande inte slutat. Nuförtiden kan en person bearbeta och använda praktiskt taget vad som helst för sina egna syften. Människan föreställer sig att vara stor, naturens kung, samtidigt som hon pekar på sina egna händers skapelser.Människan skapade allt mer perfekta redskap, först för arbete, sedan för jakt och sedan för att döda andra och deras egen sort.
För första gången insåg vi att mänskligheten är kapabel att förstöra sig själv genom att analysera konsekvenserna av en eventuell kärnvapenkonflikt. Hotet försvann och alla pustade ut. Samtidigt är explosionsenergin för alla termonukleära laddningar mindre än den energi som genereras av världens kraftverk på bara ett år. Varje år rör sig gigantiska massor av materia och förvandlas, enorma områden av den jungfruliga landytan störs, arter av växter och djur försvinner och den radioaktiva bakgrunden ökar.
Anastasia Klepneva
Mänskligheten har inte sitt ursprung i Afrika, utan i Europa. Detta sensationella uttalande gjordes av en internationell forskargrupp från University of Tübingen, Bulgarian Academy of Sciences och University of Toronto. Efter att ha studerat resterna av människoapor som hittades i Bulgarien och Grekland, kom forskare till slutsatsen att denna art av primater dök upp på europeiskt territorium för 7,2 miljoner år sedan, det vill säga minst 200 tusen år tidigare än i Afrika, som anses vara vaggan av civilisationen. RT kom på hur Bulgarien kunde bli människans förfäders hem.
- Zdenek Burian
Världens första europé
Forskare från University of Tübingen, Bulgarian Academy of Sciences och University of Toronto undersökte en tand som hittats i Bulgarien och underkäken på en apa som hittats i Grekland. Enligt experter tillhör de fossiliserade kvarlevorna människans direkta förfader, som dök upp på europeiskt territorium för cirka 7,2 miljoner år sedan - minst 200 tusen år tidigare än i Afrika. Enligt författarna till studien bevisar detta att människoapor dök upp i Europa, men sedan, på grund av ogynnsamma klimatförändringar, migrerade till den afrikanska kontinenten.
Med hjälp av datortomografi undersökte forskare den inre strukturen hos de fossiliserade resterna av Graecopithecus freybergi. I både käken och molar kunde forskare upptäcka drag som för Graecopithecus närmare apor.
"Apor tenderar att ha tydligt separerade tandrötter, och tänderna på Graecopithecus är delvis sammansmälta, vilket är karakteristiskt för moderna och antika människor, såväl som flera av deras förfäder", förklarade professor Madeline Böhme från universitetet i Tübingen, som deltog i studien.
Nytt hemland
Enligt Boehme och kollegor har klimatförändringarna tvingat människoapor att leta efter nya födokällor.
Kandidat för biologiska vetenskaper, docent vid Institutionen för antropologi, Biologiska fakulteten, Moscow State University, är dock redo att argumentera med denna hypotes. Lomonosov, vetenskaplig redaktör för portalen "Anthropogenesis.ru" Stanislav Drobyshevsky. Som han noterade kan man från afrikanska fynd spåra en direkt linje från de första aporna till moderna människor.
"Men dessa européer - ja, det här är besläktade apor, som skulle tvivla på det. De är ganska lika gorillor, men de är inte mänskliga förfäder, åtminstone enligt min personliga åsikt”, avslutade forskaren.
En tand räcker inte
Det verkar mer troligt för Stanislav Drobyshevsky att dessa antropoider dog ut i Europa. Under miocentiden, för 7-10 miljoner år sedan, levde många människoapor i både Europa och Asien. Samtidigt fanns det klimatförändringar som faktiskt tvingade dem att migrera.
"De migrerade - det är ett faktum, men det här var inte djuren som kunde lyfta och utforska en ny kontinent. Primater rör sig långsamt genom skogar. Och sen miocen var en period då skogarna försvann, så det var extremt problematiskt för dem att flytta någonstans”, förklarade Drobyshevsky. "De enda kvar är de som levde i tropiska skogar - nu har vi till exempel schimpanser och gorillor - och de som anpassat sig till den afrikanska savannen, faktiskt australopithecus." De som bodde i Europa dog lyckligtvis ut.”
Bland utländska forskare orsakar också resultaten från en internationell grupp forskare skeptiska bedömningar. Bland dem finns antropologen Peter Andrews, som var en av de första som antydde att mänskliga förfäder dök upp utanför Afrika. Han sa att att ändra sin åsikt om mänsklighetens historia baserat på bara ett fynd verkar vara ett dåligt beslut för honom.
"Uppkomsten av direkta mänskliga förfäder i Europa är i princip möjlig, men en mycket betydande mängd bevis talar för versionen av ursprunget till människan från Afrika, inklusive flera skelett och dödskallar", säger Andrews.








- Cherginets Boris Nikolaevich Historia om formation och stridsväg
- Hitlers fältmarskalkar och deras strider Jag var Hitlers adjutant Nikolaus Belov
- Hederskodex för en rysk officer i tsararmén
- Biografi om Pavel Dybenko Pavel Dybenko, ödet för revolutionens huvudseglare
- Förbindelsen mellan den första och andra västerländska armén i Smolensk År 1812 befälhavdes den västra armén av
- Så fort man blir smutsig blir den svag och äts upp
- Nürnbergrättegångar Nürnbergrättegångsprotokoll för förhör av de viktigaste krigsförbrytarna
- Konstant förändring av värdet på samma värden
- Reflektion av omsättning i redovisningsmodellen Hur man hittar varurörelsen i 1C
- skäl och stadier av övergången från batchredovisningssystem
- Avdrag från anställdas löner i 1C
- Historia om uppkomsten och utvecklingen av det uzbekiska språket
- Ring en rörmokare Miass
- Begreppet programmeringsparadigm Deklarativt och procedurminne
- Armeniska alfabetet med engelsk transkription
- Hefaistos - eldens och metallens herre
- Storskalig bekräftelse av den "afrikanska" teorin om mänskligt ursprung har erhållits
- Under vargsolen Vilken språkgrupp tillhör zigenarna?
- Uppsatser för skolbarn Tvardovsky det är inte mitt fel
- Den första ordföranden för Tunguska volost